



Содержание
Перейти к:
 Т. А. Ожаровская,
О. В. Зубкова,
О. Попова,
А. В. Ковыршина,
П. П. Голдовская,
И. В. Вавилова,
И. В. Должикова,
Е. И. Ермолова,
М. С. Кунда,
Н. Н. Рыжова,
О. Л. Воронина,
Д. Н. Щербинин,
Д. В. Щебляков,
Д. Ю. Логунов,
А. Л. Гинцбург
Т. А. Ожаровская,
О. В. Зубкова,
О. Попова,
А. В. Ковыршина,
П. П. Голдовская,
И. В. Вавилова,
И. В. Должикова,
Е. И. Ермолова,
М. С. Кунда,
Н. Н. Рыжова,
О. Л. Воронина,
Д. Н. Щербинин,
Д. В. Щебляков,
Д. Ю. Логунов,
А. Л. Гинцбург https://doi.org/10.30895/2221-996X-2024-24-3-294-311
Перейти к:
ВВЕДЕНИЕ. Вирусы Марбург и Эбола вызывают тяжелую геморрагическую лихорадку у людей и приматов. В настоящее время не зарегистрированы вакцины для одновременной профилактики вызываемых этими филовирусами заболеваний, способные предотвратить распространение заболевания или снизить его тяжесть. Исследования, направленные на выбор наиболее иммуногенной формы протективного антигена, являются необходимым этапом при разработке эффективных профилактических вакцин.
ЦЕЛЬ. Оценка индукции гуморального иммунного ответа на введение животным рекомбинантных аденовирусных векторов, экспрессирующих различные формы гликопротеина вирусов Эбола и Марбург.
МАТЕРИАЛЫ И МЕТОДЫ. Рекомбинантные аденовирусы человека 5 серотипа получали методом гомологичной рекомбинации в клетках Escherichia coli. Культивирование аденовирусов проводили в культуре клеток HEK293 с последующей очисткой методом ультрацентрифугирования в ступенчатом градиенте плотности цезия хлорида. Полученные препараты аденовирусов были охарактеризованы на подлинность методом ПЦР и полногеномным секвенированием; по количеству вирусных частиц (спектрофлуориметрия) и инфекционных вирусных частиц (метод ТЦД50). Оценку титра гликопротеин-специфических IgG антител в сыворотке крови иммунизированных животных (мыши) проводили методом иммуноферментного анализа.
РЕЗУЛЬТАТЫ. Получены рекомбинантные аденовирусы человека 5 серотипа, содержащие в своем геноме кассету с одной из форм гена гликопротеина, кодирующего полноразмерный гликопротеин (GP), GP с удаленным муциноподобным доменом, GP с удаленными гликановым кэпом и муциноподобным доменом. Каждую из форм изучали на примере GP четырех представителей филовирусов: вируса Эбола видов Заир, Судан, Бундибугио и вируса Марбург. Показано, что форма GP не оказывает критического влияния на репликативную способность рекомбинантного аденовируса. Установлено, что через 3 нед. после иммунизации наибольший уровень продукции антиген-специфичных антител вызывают аденовирусы, кодирующие полноразмерный GP и GP с удаленным муциноподобным доменом. Вне зависимости от вида филовируса антиген, представленный GP с удаленными гликановым кэпом и муциноподобным доменом, оказался наименее иммуногенным.
ВЫВОДЫ. Наиболее перспективными для разработки филовирусных вакцин на основе рекомбинантных аденовирусных векторов являются конструкции, включающие ген полноразмерного GP и GP с удаленным муциноподобным доменом.
Ожаровская Т.А., Зубкова О.В., Попова О., Ковыршина А.В., Голдовская П.П., Вавилова И.В., Должикова И.В., Ермолова Е.И., Кунда М.С., Рыжова Н.Н., Воронина О.Л., Щербинин Д.Н., Щебляков Д.В., Логунов Д.Ю., Гинцбург А.Л. Иммуногенность различных вариантов гена гликопротеина вирусов Эбола и Марбург в составе рекомбинантных аденовирусных векторов. БИОпрепараты. Профилактика, диагностика, лечение. 2024;24(3):294-311. https://doi.org/10.30895/2221-996X-2024-24-3-294-311
Ozharovskaia T.A., Zubkova O.V., Popova O., Kovyrshina A.V., Goldovskaya P.P., Vavilova I.V., Dolzhikova I.V., Ermolova E.I., Kunda M.S., Ryzhova N.N., Voronina O.L., Shcherbinin D.N., Shcheblyakov D.V., Logunov D.Y., Gintsburg A.L. Immunogenicity of various variants of Ebola and Marburg virus glycoprotein genes in recombinant adenoviral vectors. Biological Products. Prevention, Diagnosis, Treatment. 2024;24(3):294-311. (In Russ.) https://doi.org/10.30895/2221-996X-2024-24-3-294-311
EBOV — вирус Эбола вида Заир (вид Orthoebolavirus zairense);
BUDV — вирус Эбола вида Бундибугио (вид Orthoebolavirus bundibugyoense);
SUDV — вирус Эбола вида Судан (Orthoebolavirus sudanense);
MARV — вирус Марбург (вид Orthomarburgvirus marburgense);
GP — гликопротеин;
rAd5 — рекомбинантный аденовирус человека 5 серотипа;
а.о. — аминокислотный остаток;
в.ч. — вирусная частица;
ВНА — вирус-нейтрализующие антитела;
ДИ — доверительный интервал.
Вирусы Эбола и Марбург являются членами семейства Filoviridae порядка Mononegavirales и известны тем, что, будучи высоколетальными, вызывают тяжелую геморрагическую лихорадку у людей и приматов. Вирус Эбола относится к роду Orthoebolavirus, который включает 6 видов. Из них высоколетальными и патогенными для человека являются три: Orthoebolavirus zairense (EBOV), Orthoebolavirus bundibugyoense (BUDV) и Orthoebolavirus sudanense1 (SUDV). Этиологическим агентом двух наиболее крупных вспышек был вирус Эбола вида Заир. Вспышка в Западной Африке проходила в 2014–2016 гг., во время которой было инфицировано 28715 человек и зарегистрировано 11372 случая летального исхода, а вспышка в восточной части Демократической Республики Конго в 2018 г. унесла жизни еще 2320 человек2.
Поскольку летальность при инфицировании вирусом Эбола достигает 70–90%, основные усилия ученых были направлены на разработку вакцин для профилактики болезни, вызванной этим представителем семейства Filoviridae. В Российской Федерации первая подобная вакцина, названная «ГамЭвак-Комби», была разработана в ФГБУ «НИЦЭМ им. Н.Ф. Гамалеи» Минздрава России и зарегистрирована в 2015 г. (рег. удостоверение № ЛП-003390 от 28.12.2015) [1]. В 2019 г. была одобрена для медицинского применения в Европейском союзе и США вакцина ERVEBO®, разработанная учеными из Агентства общественного здравоохранения Национальной микробиологической лаборатории Канады3. В Китае была одобрена вакцина Ad5-EBOV для предотвращения заболеваний, вызванных EBOV, производства CanSino, которая продемонстрировала высокую иммуногенность и приемлемый уровень безопасности в клинических исследованиях [2].
Помимо представителей рода Orthoebolavirus, вирус Марбург, относящийся к роду Orthomarburgvirus виду marburgense (MARV), к настоящему времени явился причиной двух крупных вспышек: в Демократической Республике Конго с 1998 по 2000 гг. и в Анголе в 2004–2005 гг. (252 случая, летальность 90%) [3].
Описанные вирусы являются глобальной угрозой общественному здравоохранению из-за высокой летальности вызываемых ими заболеваний, быстрого распространения, постоянных вспышек в Западной Африке, которые могут распространиться и за пределы африканского континента, а также появления нового состояния, известного как «синдром, обусловленный перенесенной болезнью, вызванной вирусом Эбола». Заболевание проявляется в виде воспалительных и аутоиммунных реакций, таких как ревматоидный артрит, системная красная волчанка и спондилоартрит с увеитом [4].
Несмотря на то что вирус Марбург и вирус Эбола были открыты в 1967 и 1976 гг. соответственно и широко исследовались в последующие годы, в настоящее время не существует зарегистрированных вакцин для одновременной профилактики вызываемых этими филовирусами заболеваний. Показано, что протективный иммунный ответ достигается только тогда, когда антиген в составе вакцины имеет то же происхождение, что и филовирус, используемый для заражения (то есть вакцина, основанная на гликопротеине вируса Эбола вида Заир, будет обеспечивать защиту только от данного патогена). Кроме того, известно отсутствие перекрестной защиты от вирусов, антигены которых не содержатся в вакцине [5]. В связи с этим можно сделать вывод о том, что существующие вакцины могут не обеспечивать защиту от всех разновидностей филовирусов. Таким образом, филовирусы занимают одно из приоритетных мест в списке биологических угроз здравоохранению Российской Федерации.
Традиционные вакцины, такие как живые аттенуированные и инактивированные, не являются оптимальной вакцинной платформой из-за высоких рисков, связанных с недостаточной аттенуацией или неполной инактивацией летального вируса. Значительный прогресс был достигнут в использовании альтернативных платформ, таких как рекомбинантные вирусные векторы. Например, при иммунизации приматов вектором на основе аттенуированного рекомбинантного вируса везикулярного стоматита (VSV), экспрессирующего или гликопротеин (GP) EBOV, или GP MARV, животные были полностью защищены от заражения гомологичным вирусом [5].
Наиболее широко изученные векторы на основе аденовирусов (Ad) показали многообещающие результаты при создании панфиловирусной вакцины. Разработана кандидатная вакцина, состоящая из четырех рекомбинантных аденовирусов человека 5 серотипа (Ad5), каждый из которых экспрессирует по два антигена (GP пяти различных филовирусов, охватывающих три значимых вида EBOV, SUDV, MARV (три изолята) и нуклеопротеин (NP) EBOV и MARV). Иммунизация приматов продемонстрировала 100% защиту от заражения двумя видами вируса Эбола и тремя изолятами вируса Марбург, каждый из которых вводился в дозе, в 1000 раз превышающую смертельную [6].
Первую фазу клинических исследований прошла гетерологичная кандидатная вакцина, состоящая из двух компонентов. Первый компонент — Ad26.ZEBOV — представлен аденовирусом 26 серотипа, кодирующим GP EBOV, а второй — вектором на основе модифицированного вируса осповакцины Анкара, кодирующим GP EBOV, SUDV и MARV, а также NP вируса Тай Форест (названный MVA-BN-Filo) [7]. Нуклеопротеин включают в состав вакцин в качестве адъюванта, потому что известно, что NP способен вызывать эффективный клеточный иммунный ответ и может повысить эффективность вакцины на основе GP [6].
Тем не менее ключевой мишенью для разработки вакцинных препаратов является вирусный GP, который занимает центральное место в патогенности филовирусов, поскольку является ключевым структурным белком и отвечает как за проникновение вируса в клетки, так и за уклонение от иммунного ответа. Полноразмерный GP представляет собой тримерный белок слияния I типа и транслируется в белок-предшественник размером около 670 а.о., после чего расщепляется фурином клетки-хозяина на две субъединицы GP1 и GP2, связанные дисульфидной связью [8]. Субъединица GP1 отвечает за связывание с рецептором Ниманна — Пика C1 и состоит из 4 доменов: основания, головки, гликанового кэпа и муциноподобного домена [9][10]. Субъединица GP2 содержит пептид слияния и имеет наиболее высокую консервативность между различными представителями филовирусов [8]. Гликопротеин служит также основной мишенью для нейтрализующих антител. GP EBOV и MARV претерпевают сложные структурные перестройки во время проникновения вируса в клетку, что приводит к образованию различных конформационных состояний, которые могут влиять на эффективность вакцины.
Поскольку в настоящее время не созданы зарегистрированные вакцины для одновременной защиты от обоих филовирусов, способные предотвратить распространение заболевания или снизить его тяжесть, в данном исследовании мы стремились изучить иммуногенность различных форм гликопротеинов EBOV и MARV, чтобы предоставить информацию для разработки вакцин следующего поколения.
Цель работы — оценка индукции гуморального иммунного ответа на введение животным рекомбинантных аденовирусных векторов, экспрессирующих различные формы гликопротеина вирусов Эбола и Марбург.
Материалы
Бактериальные штаммы и клеточные линии
Для получения рекомбинантных плазмидных конструкций использовали лабораторные штаммы Escherichia coli DH5a (New England Biolabs, США) и BJ5183 (Stratagene, США). Для культивирования бактерий использовали ростовую среду 2xYT (Диаэм, Россия), агар бактериологический (Диаэм, Россия), селективные антибиотики ампициллин и канамицин (Thermo Fisher Scientific, США).
Для создания плазмидных конструкций, кодирующих геном rAd5 с различными формами GP вирусов рода Эбола и Марбург, использовали полученный ранее [11] экспрессионный вектор, содержащий геном rAd5 с делетированными Е1- и Е3-областями со встроенной экспрессионной кассетой.
Для получения и накопления рекомбинантных аденовирусов использовали клеточную линию HEK293 — клетки почки эмбриона человека, содержащие в геноме E1-область Ad5 (получена из Российской коллекции клеточных культур позвоночных, РККК П). Для постановки реакции нейтрализации псевдовирионов использовали клеточную линию Vero E6 — клетки почки зеленой мартышки (получена из РККК П). Клетки культивировали в среде DMEM 1х (HyClone, США) с добавлением 6% эмбриональной сыворотки крупного рогатого скота (HyClone, США), 25 мл 7,5% бикарбоната натрия (ПанЭко, Россия), 146 мг L-глутамина (ПанЭко, Россия) и смеси антибиотиков пенициллина (50 МЕ/мл) и стрептомицина (50 мкг/мл) (ПанЭко, Россия) при 37 °С в атмосфере, содержащей 5% СО2.
Лабораторные животные
Все эксперименты с использованием лабораторных животных проводили в соответствии с рекомендациями Национального стандарта Российской Федерации4. Мыши (самки возрастом 6 нед.) линии С57BL/6, экспрессирующие гаплотип b главного комплекса гистосовместимости, были получены из НПП «Питомник лабораторных животных» ФИБХ РАН (Россия). Животных содержали в системе ISOcage (Tecniplast, Италия) со свободным доступом к корму и воде. Проведение исследования было одобрено локальным независимым этическим комитетом ФГБУ «НИЦЭМ им. Н.Ф. Гамалеи» Минздрава России (выписка из протокола № 35 от 24.01.2023).
Методы
Выбор последовательностей антигена
На первом этапе работы отдельно для каждого вида филовирусов (Orthoebolavirus zairense, Orthoebolavirus bundibugyoense, Orthoebolavirus sudanense и Orthomarburgvirus marburgense) выбирали последовательность гена GP. Для этого из официального общедоступного источника NCBI Virus Variation Resource5 нами были отобраны аминокислотные последовательности полноразмерного GP штаммов филовирусов, изолированных от человека. В программе «Geneious Prime» (Biomatters, Новая Зеландия) выравнивали последовательности и получали консенсусную последовательность, в каждом положении которой находился аминокислотный остаток, наиболее часто встречающийся у полноразмерного белка. Консенсусную последовательность затем сравнивали с полными последовательностями GP изолятов данного филовируса и выбирали наиболее близкую к ней последовательность, представленную в базе данных.
Конструирование плазмидных векторов
Нуклеотидные последовательности выбранных антигенов (Zaire-GP, Zaire-GPΔM, Zaire-GPcl, Sudan-GP, Sudan-GPΔM, Sudan-GPcl, Bund-GP, Bund-GPΔM, Bund-GPcl, Marburg-GP, Marburg-GPΔM, Marburg-GPcl) были синтезированы в ЗАО «Евроген» (Россия). Полученные последовательности встраивали рестрикционно-лигазным методом клонирования в состав экспрессионной кассеты между промотором и терминатором транскрипции плазмидного вектора pSh5. Плазмидная конструкция pSh5 — это челночный вектор для гомологичной рекомбинации, несущий фрагменты генома Ad5, так называемые «плечи гомологии», а именно: левый и правый концы генома с делецией Е1-области, вместо которой встроена экспрессионная кассета (CMV-промотор, полилинкер и терминатор транскрипции (сигнал полиаденилирования вируса SV40)). Далее проводили гомологичную рекомбинацию в E. coli штамма BJ5183 между плазмидной конструкцией, несущей полный геном rAd5 с делецией Е3-области, и полученными на предыдущем этапе челночными векторами, линеаризованными по уникальному сайту рестрикции PmeI, заложенному между «плечами гомологии». Таким образом были получены 12 плазмидных конструкций, кодирующих геном rAd5 с различными формами GP вирусов рода Эбола и Марбург. Подлинность подтверждали методом ПЦР, рестрикционным картированием и полногеномным секвенированием.
Получение, накопление и очистка рекомбинантных аденовирусных векторов
Для получения рекомбинантных аденовирусов сконструированные на предыдущем этапе плазмидные конструкции обрабатывали специфической эндонуклеазой рестрикции PacI для экспонирования вирусных инвертированных концевых повторов. Полученными препаратами линейной ДНК трансфецировали клетки НЕК293 с использованием реагента Lipofectamine® 2000 (Invitrogen, США) в соответствии с рекомендациями производителя. Трансфецированные клетки пересевали с увеличением посевной площади до наступления 100% цитопатического действия. Получение рекомбинантных вирусных векторов подтверждали ПЦР и ПЦР в режиме реального времени (ПЦР-РВ). Титры аденовирусов выражали в ТЦД50/мл (50% тканевая цитопатическая доза); значения ТЦД50/мл определяли методом Рида и Менча [12].
Рекомбинантные аденовирусы нарабатывали в препаративных количествах в клетках НЕК293. Для этого 2×10⁷ клеток, высеянных на культуральные чашки диаметром 15 см, заражали рекомбинантными аденовирусами в дозе 0,5 ТЦД50 на клетку. Через 48–72 ч после трансдукции при наступлении полного цитопатического действия клетки осаждали низкоскоростным центрифугированием, ресуспендировали в фосфатно-солевом буфере и лизировали путем замораживания-оттаивания (3 цикла) для высвобождения вирусных частиц. Лизаты центрифугировали в течение 10 мин при скорости 3500 об/мин для осаждения клеточного дебриса. Супернатант концентрировали и очищали от клеточных белков и нуклеиновых кислот двукратным ультрацентрифугированием в градиенте цезия хлорида на приборе Optima™ XPN-90 в роторе SW41 (Beckman Coulter Inc., США): первое ультрацентрифугирование проводили в течение 1 ч 15 мин при 35000 об/мин и 14 °С, второе — в течение 20 ч при 41000 об/мин и 15 °С).
Концентрацию вирусных частиц в препарате определяли с помощью набора Pico488 dsDNA quantification Kit (Lumiprobe, США) согласно рекомендациям производителя. Для построения калибровочной кривой использовали стандартный образец Ad5.
Иммунизация животных и получение образцов сывороток
Для определения уровня антигенспецифического гуморального иммунного ответа мышам линии С57/BL6 (по 7 особей в группе) однократно внутримышечно вводили по 10¹° в.ч. исследуемых аденовирусных векторов (в объеме 100 мкл). Мышам в контрольной группе вводили аналогичный объем фосфатно-солевого буфера. Через 21 сут после иммунизации у мышей отбирали кровь из хвостовой вены (методом отсечения кончика хвоста) для последующего выделения сыворотки. Образцы инкубировали в течение 30 мин при температуре 37 °С, а затем центрифугировали в течение 10 мин при 300 g. Образцы сывороток хранили при минус 70 °С.
Проверка подлинности полученных рекомбинантных аденовирусов
Подлинность препаратов полученных рекомбинантных аденовирусов подтверждали методами ПЦР и ПЦР-РВ. Для этого вирусную ДНК выделяли с помощью набора Wizard Genomic DNA Purification Kit (Promega, США) согласно протоколу производителя. ПЦР-РВ проводили с использованием смеси qPCRmix-HS (ЗАО «Евроген», Россия) на приборе CFX 96 Real-Time PCR Detection System (Bio-Rad, США), детекцию проводили на канале HEX (гексахлорфлуоресцеин). Программа ПЦР-РВ представлена в таблице 1 (опубликована на сайте журнала6). Праймеры и зонд были синтезированы в ЗАО «Евроген» (Россия), их нуклеотидные последовательности представлены в таблице 2 (опубликована на сайте журнала7).
ПЦР для подтверждения наличия целевых генов проводили с использованием смеси 5X ScreenMix-HS (ЗАО «Евроген», Россия) на приборе T100™ Thermal Cycler (Bio-Rad, США). Программа амплификации представлена в таблице 3 (опубликована на сайте журнала8), нуклеотидные последовательности праймеров приведены в таблице 2 (опубликована на сайте журнала).
Секвенирование полученных аденовирусов
Библиотеки готовили по протоколу Nextera™ DNA Flex Library Prep (Illumina, США). Проверку качества и размера библиотек проводили с помощью электрофореза на чипах High Sensitivity DNA Chips 2100 Bioanalyzer System (Agilent, США). Секвенирование выполняли на платформе Illumina MiSeq c картриджем MiSeq Reagent Kit v2 Nano (500 Cycles). Для сборки прочтений de novo и по предоставленным референсным последовательностям использовали пакет программ CLC Genomic Workbench v.21 (Qiagen, США).
Определение титра антител в сыворотке крови методом иммуноферментного анализа
Титр GP-специфических антител в сыворотке крови иммунизированных животных определяли методом иммуноферментного анализа по стандартному протоколу. Использовали следующие рекомбинантные белки: GP вируса Марбург с делецией трансмембранного домена (MARV-Angola rGPΔTM) (IBT Bioservices, США); полный GP вируса Эбола вида Sudan штамма Gulu (SinoBiological, КНР); GP1-домен гликопротеина вируса Эбола вида Bundibugyo штамма Uganda 2007 GP1/Glycoprotein Protein (His Tag), (SinoBiological, КНР); полный GP вируса Эбола вида Zaire штамма H. sapiens-wt/GIN/2014/Kissidougou-C15 (SinoBiological, КНР). Для детекции использовали вторичные антитела, специфичные к IgG мыши и конъюгированные с пероксидазой хрена (IgG-NXA931, GE Healthсare, США). Оптическую плотность измеряли при длине волны 450 нм на планшетном спектрофотометре Multiscan™ FC (Thermo Fisher Scientific, США). За титр антител принимали обратную величину последнего разведения сыворотки крови при условии, что значение оптической плотности целевого образца превосходит таковое контрольного образца >2 раза.
Реакция нейтрализации псевдовирионов
Для постановки анализа четыре рекомбинантных VSV (rVSV-Zaire-GP, rVSV-Sudan-GP, rVSV-Bund-GP и rVSV-Marburg-GP (псевдотипированные GP вируса Эбола или Марбург)) получали, как описано ранее [13]. Для проведения эксперимента клетки линии Vero E6 (10⁶ клеток на лунку) высевали на 6-луночные планшеты за день до эксперимента. Перед проведением эксперимента сыворотки крови мышей инактивировали при температуре 56 °С в течение 30 мин. Двукратные разведения образцов инактивированных сывороток мышей (начиная с 1:10) смешивали с rVSV-GP в количестве 100 бляшкообразующих единиц, а затем полученную смесь инкубировали в течение 1 ч при 37 °C. Далее смесь добавляли к монослою клеток Vero Е6 в трех повторах. Через 6 ч отбирали среду, наносили полутвердое агаризированное покрытие на клетки и инкубировали в течение 48 ч при 37 °С в атмосфере, содержащей 5% CO2. После этого проводили учет результатов. За титр нейтрализующих антител в сыворотке крови иммунизированного животного принимали обратную величину максимального разведения, при котором наблюдали 50% снижение количества бляшек rVSV-GP по сравнению с сыворотками не вакцинированных животных.
Определение титра вирус-нейтрализующих антител к Ad5 в сыворотках крови иммунизированных животных
Титры вирус-нейтрализующих антител (ВНА) к вектору Ad5 измеряли с помощью реакции вирус-нейтрализации на культуре клеток HEK293. Для анализа использовали Ad5, экспрессирующий ген зеленого флуоресцентного белка (Ad5-EGFP) [14]. Двукратные разведения образцов сывороток (начиная с 1:25) вносили в лунки 96-луночного планшета. К ним добавляли по 50 мкл Ad5-EGFP (в дозе 100 ТЦД50) и инкубировали в течение 1 ч при 37 °С. Каждую сыворотку анализировали в трех повторах. Далее в каждую лунку планшета вносили по 100 мкл клеточной суспензии (5×10⁶ клеток). Планшеты инкубировали в течение 48 ч при 37 °С в атмосфере, содержащей 5% CO2. Флуоресценцию ЕGFP определяли при помощи инвертированного микроскопа Olympus IX73 (Olympus; Япония) с модулем U-RFL-T (Olympus; Япония) при длинах волн 490 и 510 нм возбуждения и эмиссии соответственно. Титры ВНА определяли количественно как обратную величину разведения сыворотки крови мышей в лунке, где достигалось снижение количества фокусов флуоресценции на 50% по сравнению с лункой положительного контрольного образца (содержащего только Ad5-EGFP без добавления сыворотки).
Статистический анализ
Статистический анализ проводили при помощи программы GraphPad Prism 8.0.1 (GraphPad Software, США), а также Exсel (Microsoft, США). Для анализа несвязанных выборок применяли критерий Манна–Уитни. Результаты сравнения экспериментальных и контрольных групп считали статистически достоверными при значении p<0,05.
Выбор последовательностей и различных форм гликопротеинов вирусов Эбола и Марбург
На первом этапе работы осуществляли выбор последовательностей антигенов четырех филовирусов. Поскольку известно, что вирус EBOV обладает достаточно высокой скоростью мутирования (скорость замен составляет 7,06×10-⁴ нуклеотидных замен / позицию нуклеотида / год) [15], для выбора последовательности GP вируса EBOV использовали последовательности изолятов вируса Эбола (наиболее крупной вспышки (2014–2016 гг.), унесшей жизни 11372 человек). Консенсусная последовательность совпала с последовательностью GP GenBank: AIG95977.
Несмотря на то что вирусы EBOV и SUDV впервые были обнаружены в один и тот же год (1976 г.), установлено, что вирус SUDV характеризуется в 15 раз меньшей скоростью мутирования (0,46×10–⁴ нуклеотидных замен / позицию нуклеотида / год). Поэтому для выбора последовательности GP вируса SUDV использовали консенсусные последовательности изолятов, построенные по всем доступным в базе данных NCBI Virus Variation Resource9 последовательностям (в том числе по последовательностям изолятов, полученных в ходе пяти крупных вспышек заболевания); результирующая последовательность совпала с GP GenBank: AGB56678. Вирусы BUDV и MARV характеризуются более низкой мутационной изменчивостью, поэтому в качестве целевых были выбраны консенсусные последовательности, которые наиболее гомологичны последовательностям GP циркулирующих изолятов (GenBank: AGL73460 и GenBank: AFV31307 соответственно).
Затем были разработаны модификации гена GP для каждого вируса (рис. 1). В белке GPΔM отсутствует полный Ser/Thr муциноподобный домен, который включает большую С-концевую часть субъединицы GP1, обогащенную сайтами N- и O-гликозилирования [16]. Поскольку гликаны вирусного белка играют решающую роль в уклонении от иммунного ответа хозяина, образуя так называемый «гликановый кэп» на поверхности GP, то для сравнения нами была выбрана последовательность (названная «GPcl»), в которой удалены как муциноподобный домен, так и гликановый кэп — наиболее гликозилированные области данного антигена.
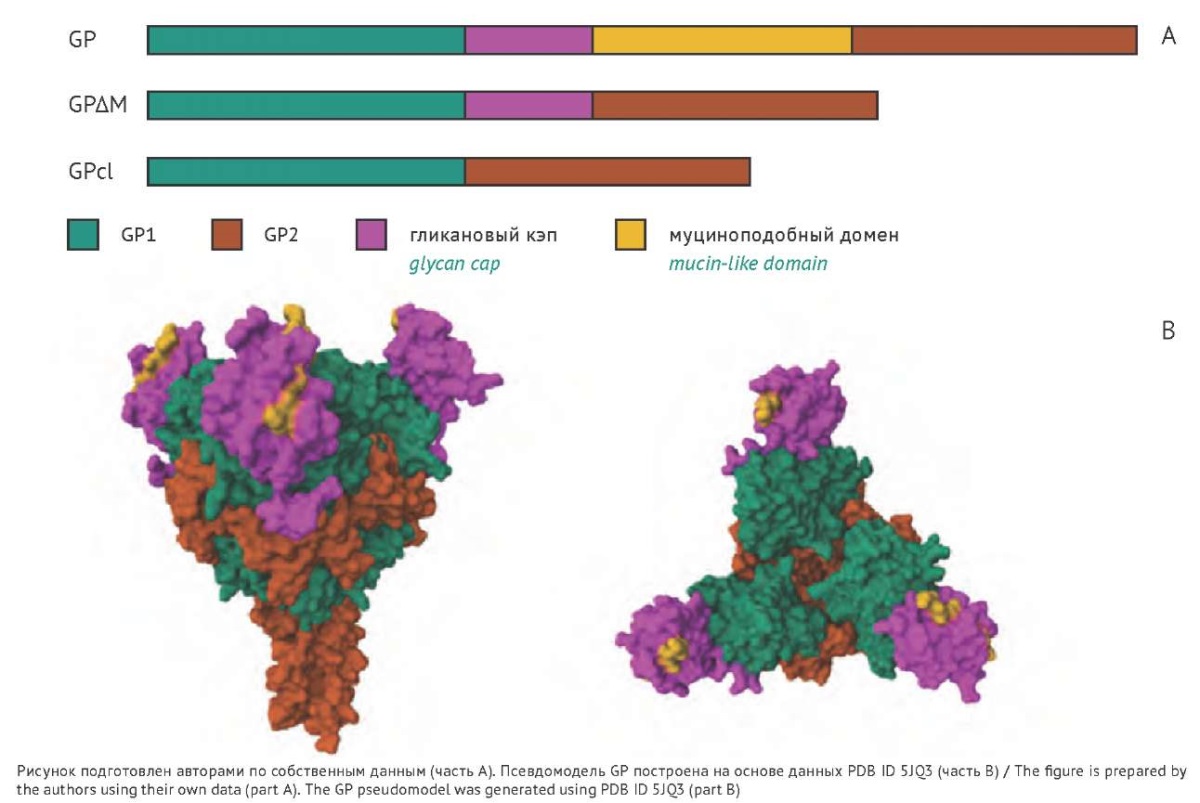
Рис. 1. Схематичное изображение последовательности и структуры гликопротеина (GP) филовируса: А — схема доменной структуры полноразмерного гена GP филовируса и исследуемых модифицированных форм (GPΔM и GPcl); B — псевдомодель GP филовируса с обозначением тех же доменов. Модель построена с помощью RCSB PDB на основе PDB ID 5JQ3 [17]. Желтым цветом обозначено приблизительное расположение муциноподобного домена, соединяющего соответствующие концы гликанового кэпа (фиолетовый цвет) и субъединицы GP2 (оранжевый цвет). Часть муциноподобного домена GP, которая считается неструктурированной, не представлена на модели.
Fig. 1. Schematic representation of the filovirus glycoprotein (GP) sequence and structure. A, schematic domain structure of the full-length form (GP) and the studied modified forms (GPΔM and GPcl) of the filovirus GP gene. B, filovirus GP pseudomodel with the same domains highlighted in the same colours. The yellow colour indicates the approximate location of the mucin-like domain connecting the corresponding ends of the glycan cap (magenta) and the GP2 subunit (orange). The model does not show the GP mucin-like domain part that is considered unstructured. The model was generated using the PDB ID 5JQ3 entry from the Protein Data Bank of the Research Collaboratory for Structural Bioinformatics (RCSB PDB) [17].
Получение рекомбинантных аденовирусов, кодирующих различные формы гликопротеинов филовирусов
Для определения и сравнения иммуногенности различных форм GP панфиловирусов нами было получено двенадцать rAd5, содержащих на месте удаленной Е1-области своего генома экспрессионную кассету с геном одной из форм GP, в соответствии со стандартными протоколами для создания аденовирусных векторов. Схемы всех полученных рекомбинантных аденовирусов показаны на рисунке 2.
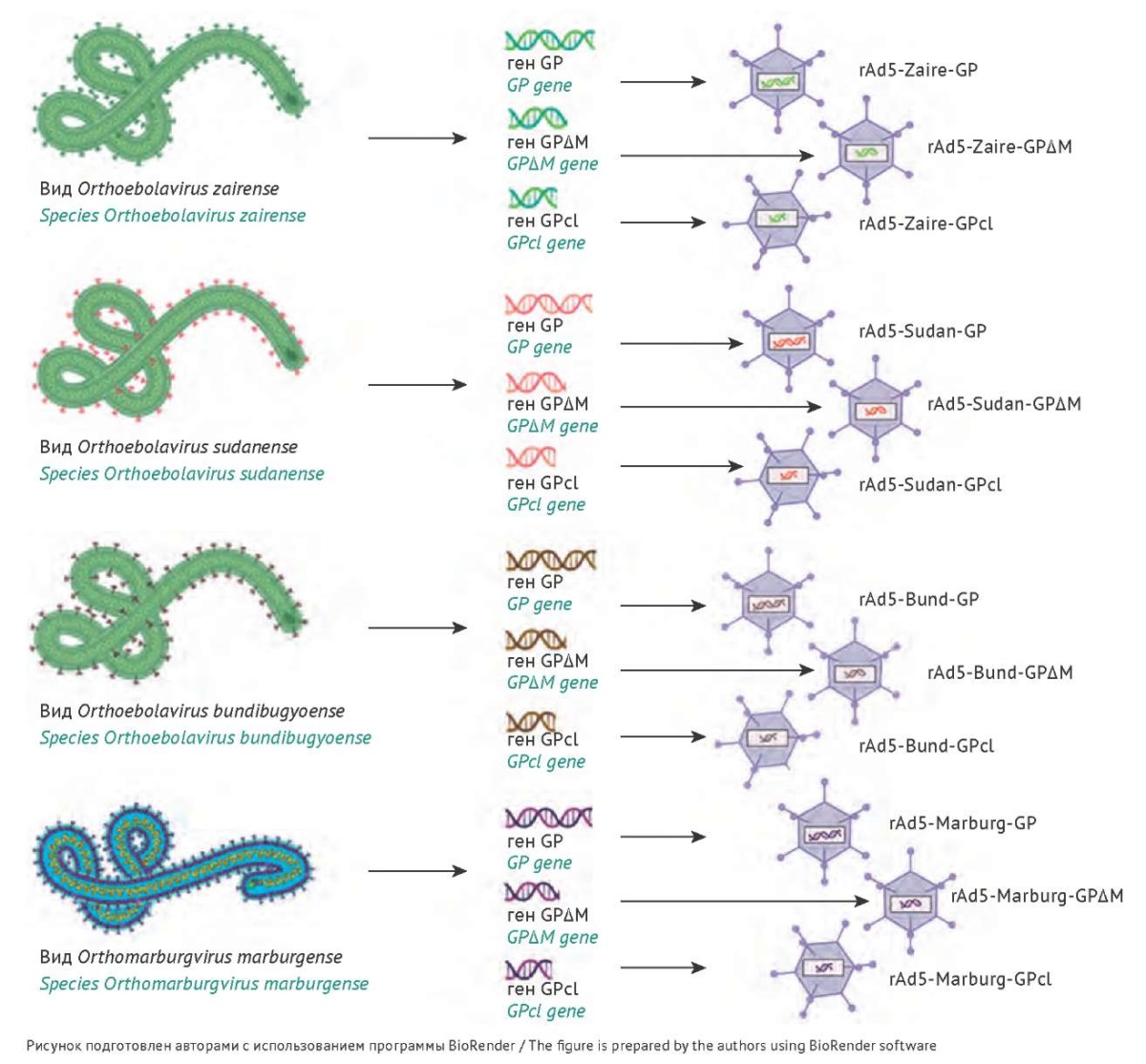
Рис. 2. Схематичное изображение рекомбинантных аденовирусов (rAd5), содержащих в геноме кассету с одной из форм гликопротеина (GP) соответствующего филовируса.
Fig. 2. Schematic representation of recombinant adenoviruses (rAd5), each containing a cassette with one of the glycoprotein (GP) forms of the corresponding filovirus.
В настоящее время технология получения rAd5 хорошо изучена и отработана. Однако наряду с относительной простотой создания rAd5 существует сложность лежащих в основе данного процесса биологических характеристик самого вируса: можно упустить из виду возможность того, что культуральные свойства рекомбинантного аденовируса будут снижены за счет гиперэкспрессии целевого трансгена. Каждая новая комбинация вектора и целевого гена может рассматриваться как новая система, подвергающаяся искусственному отбору в лабораторной культуре клеток. В связи с этим нами были изучены культуральные свойства всех полученных rAd5. Для этого вирусы наращивали на десяти культуральных чашках, проводили очистку методом ультрацентрифугирования и измеряли количество в.ч., определяли инфекционный титр и число геномов rAd5 (табл. 4) (опубликована на сайте журнала10). По результатам исследований во всех пробах был обнаружен ген гексона rAd5. Соотношение в.ч./ТЦД50 находилось в диапазоне значений от 500 до 1000 (стандартный диапазон при культивировании rAd5).
Установлено, что удаление муциноподобного домена и гликанового кэпа GP не оказывает влияния на рост rAd5, несущего ген GP вируса Марбург. При одинаковых условиях культивирования среди вирусов, кодирующих GP EBOV, наибольшее количество в.ч. после очистки наблюдалось у rAd5-Zaire-GPcl. При этом среди rAd5, кодирующих различные формы GP SUDV и BUDV, аденовирусы, несущие ген полноразмерного белка GP, были накоплены в большем количестве. Представленные в таблице 4 (опубликована на сайте журнала11) данные показывают, что форма GP не оказывает критического влияния на выход рекомбинантного аденовируса. В тех случаях, когда экспрессия трансгена препятствует размножению аденовирусного вектора, существует возможность получения векторов с мутациями в целевом гене, что обеспечивает улучшение репликативных свойств по сравнению с исходным вирусом [18].
Согласно полученным данным ПЦР и полногеномного секвенирования подлинность всех полученных аденовирусов была подтверждена (данные не представлены).
Сравнение иммуногенности различных форм гликопротеинов вирусов Эбола и Марбург в составе аденовирусных векторов
Для анализа иммуногенных свойств различных форм GP мышам внутримышечно однократно вводили по 100 мкл каждого из препаратов полученных 12 рекомбинантных аденовирусов в дозе 10¹° в.ч. Через 3 нед. после иммунизации получали образцы сывороток крови животных и анализировали в них титры антигенспецифических антител. Индивидуальные данные, а также среднее геометрическое значение титра (СГТ), представлены на рисунке 3. В сыворотке крови мышей контрольной группы (животные, которым вводили фосфатно-солевой буфер) GP-специфические IgG не обнаружили.
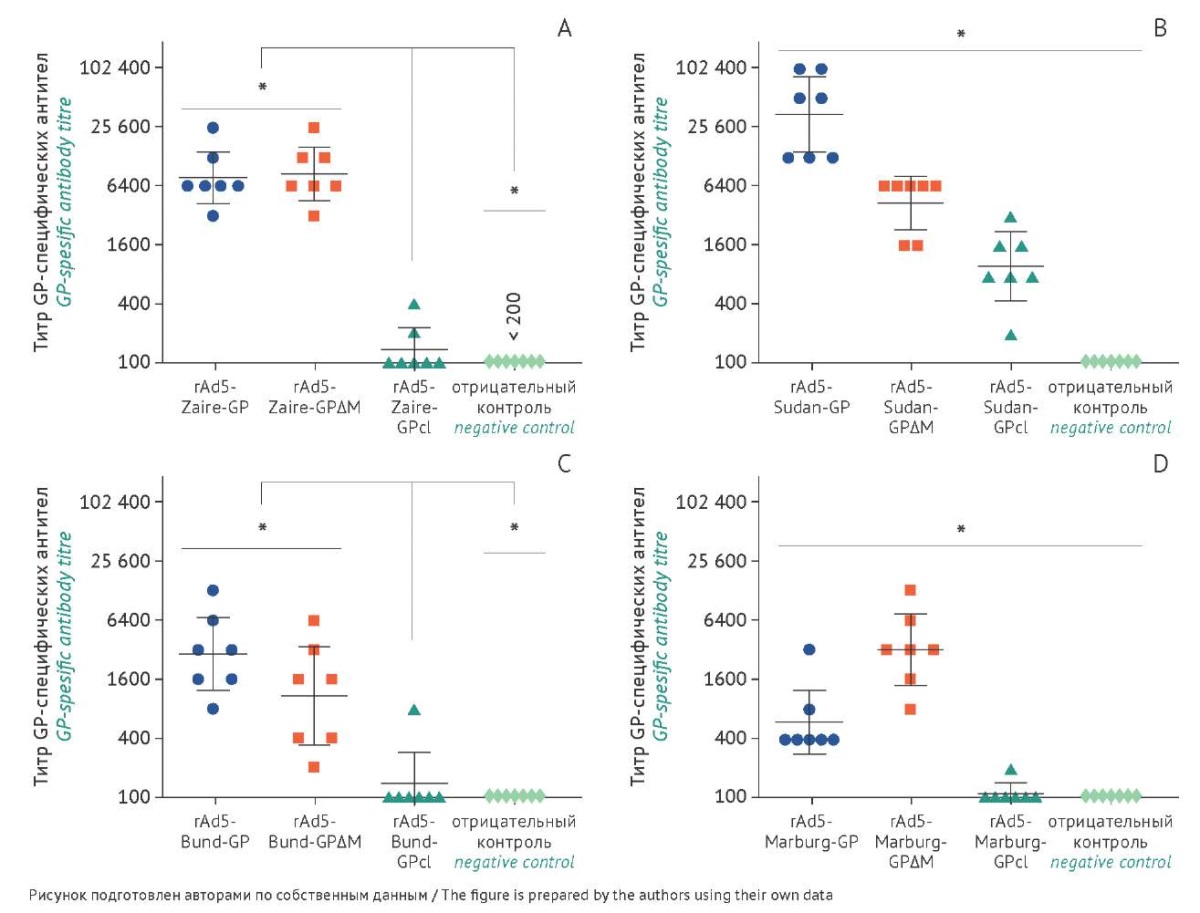
Рис. 3. Иммуногенность рекомбинантных аденовирусов (rAd5), кодирующих различные формы гликопротеина (GP) филовирусов. Данные представлены в виде индивидуальных значений титров GP-специфических антител в сыворотках крови иммунизированных животных. По оси Х обозначены группы мышей, иммунизированных векторами, экспрессирующими различные формы GP EBOV (А), SUDV (В), BUDV (С), MARV (D); отрицательный контроль — животные, которым вводили фосфатно-солевой буфер. По оси Y — индивидуальные значения титров GP-специфических антител. Отмечен средний геометрический титр (СГТ) и 95% доверительный интервал СГТ для каждой группы (n=7). A — титры антител, специфичных к GP EBOV (* — p<0,05, отличия статистически значимы при сравнении rAd5-Zaire-GPcl с остальными группами). B — титры антител, специфичных к GP SUDV (* — p<0,05, отличия статистически значимы между всеми группами). C — титры антител, специфичных к GP BUDV (* — p<0,05, отличия статистически значимы при сравнении rAd5-Bund-GPcl с остальными группами). D — титры антител, специфичных к GP MARV (* — p<0,05, отличия статистически значимы между всеми группами).
Fig. 3. Immunogenicity of recombinant adenoviruses (rAd5) encoding various filovirus glycoprotein (GP) forms. Data are presented as individual values of GP-specific antibody titres in the sera of immunised animals. The X-axis shows groups of mice immunised with vectors expressing different GP forms of Zaire Ebola virus (EBOV; A), Sudan Ebola virus (SUDV; B), Bundibugyo Ebola virus (BUDV; C), and Marburg virus (MARV; D), as well as negative control animals injected with phosphate-buffered saline. The Y-axis shows individual values of GP-specific antibody titres. The geometric mean titre (GMT) and the 95% confidence interval are indicated for each group (n=7). A, EBOV GP-specific antibody titres (* p<0.05; the differences between rAd5-Zaire-GPcl and the other groups are statistically significant). B, SUDV GP-specific antibody titres (* p<0.05; the differences between all groups are statistically significant). C, BUDV GP-specific antibody titres (* p<0.05; the differences between rAd5-Bund-GPcl and the other groups are statistically significant). D, MARV GP-specific antibody titres (* p<0.05; the differences between all groups are statistically significant).
Наибольший титр антител, специфичных к GP EBOV, наблюдали в группе животных, иммунизированных rAd5-Zaire-GPΔM (СГТ 8614, 95% доверительный интервал (ДИ) 4608–16103), чуть ниже — в группе rAd5-Zaire-GP (СГТ 7802, 95% ДИ 4240–14355), однако данная разница не была статистически достоверна (p=0,8491, критерий Манна–Уитни). В группе животных, иммунизированных rAd5-Zaire-GPcl, GP-специфические IgG наблюдали только у 2 из 7 мышей.
По результатам проведенного исследования показано, что все три полученных рекомбинантных аденовируса, несущих ген GP SUDV, индуцируют формирование специфичного гуморального иммунного ответа. При этом наибольший титр антител наблюдали в группе животных, иммунизированных rAd5-Sudan-GP (СГТ 34455, 95% ДИ 14068–84385). У животных, иммунизированных rAd5-Sudan-GPΔM, обнаружено достоверное снижение титров антител (СГТ 4307, 95% ДИ 2304-8051) по сравнению с rAd5-Sudan-GP, а самые низкие титры отмечены у мышей, которым вводили rAd5-Sudan-GPcl (СГТ 975, 95% ДИ 437–2178).
Наибольший титр GP-специфических IgG наблюдали в группе животных, иммунизированных rAd5-Bund-GP (СГТ 2898, 95% ДИ 1224–6865). У мышей, иммунизированных rAd5-Bund-GPΔM, титры антител (СГТ 1077, 95% ДИ 337–3442) статистически значимо не отличались от группы rAd5-Bund-GP. У животных, иммунизированных rAd5-Bund-GPcl, специфические антитела обнаружили только у одной мыши.
Наибольший титр антител, специфичных к GP MARV, наблюдали в группе животных, иммунизированных rAd5-Marburg-GPΔM (СГТ 3200, 95% ДИ 1399–7321), при этом титры антител в группе rAd5-Marburg-GP (СГТ 594, 95% ДИ 287–1230) были достоверно ниже (p=0,0064, критерий Манна–Уитни). В группе животных, иммунизированных Ad5-Marburg-GPcl, GP-специфические IgG наблюдали только у 1 из 7 мышей.
Отметим, что форма GP с удаленными муциноподобным доменом и гликановым кэпом (GPcl) оказалась наименее иммуногенной вне зависимости от вида филовируса. Возможно, такой белок не приобрел благоприятную конформацию, необходимую для обеспечения адекватного представления иммунной системе животных и выработки сильного гуморального ответа у мышей. Однако известно, что в области гликанового кэпа находятся два иммунодоминантных CD4+ Т-клеточных эпитопа (а.о. в положениях 244–263 и 252–271) [19], потеря которых снижает как гуморальный, так и клеточный иммунный ответ. Поскольку CD4+ Т-клетки необходимы для индукции устойчивости к заражению вирусом Эбола [19], а гликановый кэп, по-видимому, имеет решающее значение для обеспечения защитного иммунного ответа у мышей против летальной дозы EBOV [20], представленные авторами в данной статье результаты подтверждают значимость муциноподобного домена и гликанового кэпа для эффективного иммунного ответа.
Ранее было показано, что использование в качестве антигена полноразмерного GP EBOV вызывает появление антител, нацеленных преимущественно на гликановый кэп и муциноподобный домен [21][22]. Данные антитела, как правило, не обладают нейтрализующей активностью, поскольку оба домена отщепляются во время проникновения EBOV в клетки до того, как произойдет связывание с рецептором и слияние [23]. Однако позднее было описано несколько вариантов антител, связывающихся с эпитопами на гликановом кэпе, которые эффективно нейтрализовали EBOV и являлись полифункциональными (то есть способными выполнять несколько функций одновременно, в отличие от монофункциональных антител, которые могут связываться только с одним специфическим антигеном или выполнять одну задачу) [24][25].
Полученные нами результаты демонстрируют важность использования полноразмерного GP или GP с удаленным муциноподобным доменом для индукции GP-специфических IgG. Дополнительная делеция гликанового кэпа практически лишает GP иммуногенных свойств.
Исследование перекрестной реактивности антител к гликопротеинам вирусов Эбола и Марбург
Аминокислотные последовательности гликопротеинов BUDV, EBOV и SUDV различаются >30%, а отличия с GP вируса MARV составляют >70% [26]. Поэтому нами было проведено исследование перекрестной реактивности антител к GP вирусов Эбола и Марбург. Образцы сывороток крови животных, полученные в предыдущем эксперименте, были пулированы и проанализированы на наличие перекрестно реагирующих GP-специфических антител (рис. 4). Из исследования были исключены группы животных, которым вводили аденовирусы, экспрессирующие ген GP с удаленными муциноподобным доменом и гликановым кэпом (GPcl), из-за их низкой иммуногенности.
Рис. 4. Титры антител класса IgG, специфичных к гликопротеину (GP) вируса Эбола вида Заир (EBOV) — А, Судан (SUDV) — B, Бундибугио (BUDV) — C и вируса Марбург (MARV) — D, в сыворотках крови мышей, иммунизированных рекомбинантными аденовирусными векторами (rAd5). По оси Х обозначены группы мышей, иммунизированных векторами, экспрессирующими различные формы GP EBOV (А), SUDV (В), BUDV (С), MARV (D); отрицательный контроль — животные, которым вводили фосфатно-солевой буфер. По оси Y — среднегеометрический титр (СГТ) GP-специфических антител. Данные представлены в виде СГТ пулированных сывороток, исследованных в четырех повторах; отмечено стандартное отклонение.
Fig. 4. Titres of IgG antibodies specific to glycoprotein (GP) forms of Zaire Ebola virus (EBOV; A), Sudan Ebola virus (SUDV; B), Bundibugyo Ebola virus (BUDV; C), and Marburg virus (MARV; D) in the sera of mice immunised with recombinant adenoviral vectors (rAd5). The X-axis shows groups of mice immunised with vectors expressing different GP forms of EBOV (A), SUDV (B), BUDV (C), and MARV (D), as well as negative control animals injected with phosphate-buffered saline. The Y-axis shows geometric mean titre values (GMT) for GP-specific antibodies. Data are presented as GMT values for pooled sera tested in quadruplicate; the standard deviation is indicated.
Анализ результатов проведенного исследования указал на то, что иммунизация мышей рекомбинантными аденовирусами, экспрессирующими две формы гена GP SUDV (rAd5-Sudan-GP и rAd5-Sudan-GPΔM), приводит к формированию перекрестного гуморального иммунитета к GP EBOV и BUDV. Кроме того, GP-специфические антитела к EBOV, SUDV и BUDV не обладают перекрестной активностью к GP MARV. В свою очередь, антитела в сыворотке крови мышей, иммунизированных rAd5-Marburg-GPΔM, обладают перекрестной активностью к GP EBOV и SUDV и определяются в минимальных титрах (СГТ составили 100 и 71 соответственно).
Кроме того, была детектирована перекрестная реактивность антител, специфичных к EBOV и SUDV, а также к GP BUDV. Это согласуется с проведенным ранее исследованием, где была показана как перекрестная реактивность, так и защита от заражения летальной дозой BUDV после иммунизации кандидатными векторными вакцинами, экспрессирующими GP EBOV и SUDV [27].
Определение вирус-нейтрализующих антител к гликопротеину филовирусов в сыворотках крови иммунизированных животных
Наличие нейтрализующих антител у обезьян не коррелирует с защитой от инфекции, вызванной вирусом Эбола. Кроме того, нейтрализующие антитела у вакцинированных животных могут не обнаруживаться [27]. Тем не менее для более полного сравнения иммуногенности различных форм GP нами было проведено исследование индукции вирус-нейтрализующих антител (ВНА) после иммунизации животных.
При традиционной постановке эксперимента по определению ВНА к вирусам Эбола и Марбург требуется использование инфекционного вируса, обращение с которым должно осуществляться в лабораториях соответствующего уровня биобезопасности, поскольку все филовирусы относятся к микроорганизмам I группы патогенности (уровень биобезопасности 4 (BSL-4)). В связи со сложными условиями постановки такого эксперимента, трудоемкости и времязатратности (до 4 сут) было принято решение использовать аналогичный анализ — анализ нейтрализации псевдовирионов (PVNA). В качестве псевдовируса был выбран VSV, поскольку с данным вирусом можно работать в лабораториях уровня BSL-2, а сам тест является безопасным, быстрым (48 ч) и масштабируемым.
Для анализа использовали предварительно инактивированные пулированные сыворотки крови мышей (из анализа исключили группы животных, которым вводили rAd5, экспрессирующие ген GPcl из-за их низкой иммуногенности). Наибольший титр ВНА наблюдали в группах животных, иммунизированных rAd5-Marburg-GP и rAd5-Marburg-GPΔM (СГТ составило 40) (рис. 5). Титр ВНА к SUDV отличался между группами: у мышей, иммунизированных rAd5-Sudan-GP, СГТ составил 40, и rAd5-Sudan-GPΔM — СГТ составил 10. Титр ВНА к EBOV оказался минимальным (СГТ — 10), а ВНА к BUDV у животных, иммунизированных аденовирусами rAd5-Bund-GP и rAd5-Bund-GPΔM, обнаружены не были.
Рис. 5. Титры вирус-нейтрализующих антител (ВНА), специфичных к EBOV GP (А), SUDV (B), BUDV — (C) и MARV — (D), в сыворотках крови мышей, иммунизированных rAd5. По оси Х обозначены группы мышей, иммунизированных векторами, экспрессирующими различные формы GP EBOV (А), SUDV (В), BUDV (С), MARV (D); отрицательный контроль — животные, которым вводили фосфатно-солевой буфер. По оси Y — среднегеометрический титр (СГТ) ВНА. Данные представлены в виде СГТ пулированных сывороток, исследованных в трех повторах; отмечено стандартное отклонение.
Fig. 5. Titres of virus-neutralising antibodies (VNAs) specific to glycoprotein (GP) forms of Zaire Ebola virus (EBOV; A), Sudan Ebola virus (SUDV; B), Bundibugyo Ebola virus (BUDV; C), and Marburg virus (MARV; D) in the sera of mice immunised with recombinant adenoviral vectors (rAd5). The X-axis shows groups of mice immunised with vectors expressing different GP forms of EBOV (A), SUDV (B), BUDV (C), and MARV (D), as well as negative control animals injected with phosphate-buffered saline. The Y-axis shows geometric mean titre values (GMT) for VNAs. Data are presented as GMT values for pooled sera tested in quadruplicate; the standard deviation is indicated.
Полученные результаты согласуются с предыдущими исследованиями, в которых показано, что ВНА не всегда детектируются после вакцинации, однако животные выживают после заражения летальной дозой филовируса [5]. Так, титры ВНА к EBOV у макак-крабоедов (Macaca fascicularis), иммунизированных рекомбинантным VSV, экспрессирующим GP SUDV, не были обнаружены, несмотря на высокие титры IgG (конечные разведения от 500 до 10000). Однако ВНА стали детектироваться (конечные разведения от 80 до 320) только через 14 и 28 сут после заражения летальной дозой вируса [5]. В том же исследовании у макак-крабоедов, иммунизированных рекомбинантным VSV, экспрессирующим GP MARV, ВНА были обнаружены только у 1 из 4 обезьян, несмотря на наличие антител класса IgG и протективных свойств данного вектора против летальной дозы вируса Марбург [5]. В другом исследовании показано, что у яванских макак, иммунизированных репликонами альфа-вируса, кодирующими GP MARV, не были детектированы ВНА к вирусу Марбург, однако эти животные выживали после заражения летальной дозой вируса [28]. Это позволяет предположить, что появление протективного иммунитета может являться результатом довольно высоких уровней ненейтрализующих антител (от англ. non-neutralising antibodies) у животных.
Определение вирус-нейтрализующих антител к вектору в сыворотках крови иммунизированных животных
Помимо ВНА к целевому белку, при иммунизации происходит индукция ВНА к вектору, если он обладает вирусной природой. Для определения уровня ВНА была поставлена реакция нейтрализации вирусов с образцами пулированных сывороток (рис. 6). В результате титры ВНА к Ad5 были низкими (СГТ<100) во всех образцах.
Рис. 6. Титры вирус-нейтрализующих антител (ВНА) к Ad5 в сыворотках крови мышей, иммунизированных рекомбинантными аденовирусными векторами (rAd5), кодирующими различные формы GP. По оси Х обозначены группы мышей, иммунизированных векторами, экспрессирующими различные формы GP EBOV, SUDV, BUDV, MARV; отрицательный контроль — животные, которым вводили фосфатно-солевой буфер. По оси Y — среднегеометрический титр (СГТ) ВНА. Данные представлены в виде СГТ пулированных сывороток, исследованных в четырех повторах; отмечено стандартное отклонение.
Fig. 6. Titres of virus-neutralising antibodies (VNAs) against adenovirus type 5 (Ad5) in the sera of mice immunised with recombinant adenoviral vectors (rAd5) encoding various glycoprotein (GP) forms. The X-axis shows groups of mice immunised with vectors expressing different GP forms of Zaire Ebola virus (EBOV), Sudan Ebola virus (SUDV), Bundibugyo Ebola virus (BUDV), and Marburg virus (MARV), as well as negative control animals injected with phosphate-buffered saline. The Y-axis shows geometric mean titre values (GMT) for VNAs. Data are presented as GMT values for pooled sera tested in quadruplicate; the standard deviation is indicated.
В настоящее время среди исследователей нет единого мнения по поводу пороговых значений титров ВНА к Ad5 в реакции нейтрализации вирусов. В работе F-C. Zhu с соавт. [29] предложено считать титр 1:200 в сыворотках крови человека в качестве точки отсечения между низкими и высокими титрами ВНА к Ad5. В других исследованиях мышей иммунизировали однократно и двукратно Ad5 в дозе 10¹° в.ч. для создания предсуществующего иммунитета к вектору. В результате титры ВНА к Ad5 через 4 нед. составили около 1:20 при однократной и более 1:200 при двукратной иммунизации [30, 31]. В качестве порогового значения высокого титра ВНА также принят титр 1:200. В результате нами было установлено, что при однократной иммунизации мышей индуцируется образование ВНА к вакцинному вектору только в низких титрах (СГТ <100).
Одним из направлений дальнейших исследований является изучение полученных векторов на других видах животных, поскольку существуют как высокоиммунодоминантные эпитопы GP, которые распознаются антителами в поликлональных сыворотках крови (смесь антител, выработанных различными клонами В-лимфоцитов в ответ на антигенное воздействие, с широким спектром специфичности к различным эпитопам целевого антигена) нескольких разных видов животных одновременно (мышей, морских свинок и макак-резус), так и эпитопы, которые могут чаще распознаваться сывороточными антителами, полученными от конкретных видов животных [32].
Таким образом, при разработке векторных вакцин для профилактики филовирусных инфекций в качестве антигена можно использовать как полноразмерный GP, так и GP с удаленным муциноподобным доменом всех четырех филовирусов: вируса Эбола вида Заир, Судан, Бундибугио и вируса Марбург.
Вклад авторов. Все авторы подтверждают соответствие своего авторства критериям ICMJE. Наибольший вклад распределен следующим образом: Т.А. Ожаровская — участие в формулировании концепции и плана экспериментов, получение генно-инженерных конструкций, получение и наработка рекомбинантных аденовирусов, оценка гуморального иммунного ответа, исследование вирус-нейтрализующих антител к аденовирусу, написание текста рукописи; О.В. Зубкова — формулирование концепции и плана экспериментов, получение рекомбинантных аденовирусов, исследование вирус-нейтрализующих антител к аденовирусу, формулировка выводов, редактирование рукописи; О. Попова — получение генно-инженерных конструкций, оценка гуморального иммунного ответа, постановка реакции нейтрализации псевдовирусов; А.В. Ковыршина, И.В. Вавилова — анализ препаратов рекомбинантных аденовирусов; П.П. Голдовская — комплекс работ по экспериментам с животными; И.В. Должикова — участие в формулировании концепции и плана экспериментов, выбор последовательностей антигенов; Е.И. Ермолова, М.С. Кунда, Н.Н. Рыжова, О.Л. Воронина — секвенирование рекомбинантных аденовирусов; Д.Н. Щербинин — биоинформатический анализ и выбор последовательностей антигенов; Д.В. Щебляков — редактирование текста рукописи; Д.Ю. Логунов, А.Л. Гинцбург — утверждение окончательной версии статьи для публикации.
Соответствие принципам этики. Исследование было одобрено на заседании этического комитета ФГБУ «НИЦЭМ им. Н.Ф. Гамалеи» Минздрава России, протокол заседания № 35 от 24.01.2023.
Дополнительная информация. На сайте журнала «БИОпрепараты. Профилактика, диагностика, лечение» размещены таблицы 1–4.
https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table1
https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table2
https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table3
https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table4
Authors’ contributions. All the authors confirm that they meet the ICMJE criteria for authorship. The most significant contributions were as follows. T.A. Ozharovskaia participated in the formulation of the concept and plan of experiments, obtained genetically engineered constructs, obtained and grew recombinant adenoviruses, assessed the humoral immune response and adenovirus-neutralising antibodies, and drafted the manuscript. O.V. Zubkova formulated the concept and plan of experiments, produced recombinant adenoviruses, studied adenovirus-neutralising antibodies, formulated the conclusions, and edited the manuscript. O. Popova obtained genetically engineered constructs, assessed the humoral immune response, and conducted pseudovirus neutralisation tests. A.V. Kovyrshina and I.V. Vavilova analysed the recombinant adenovirus samples. P.P. Goldovskaya conducted the experiments with animals. I.V. Dolzhikova participated in the formulation of the concept and the plan of experiments and selected antigen sequences. E.I. Ermolova, M.S. Kunda, N.N. Ryzhova and O.L. Voronina performed sequencing of recombinant adenoviruses. D.N. Shcherbinin conducted bioinformatics analysis selected antigen sequences. D.V. Shcheblyakov edited the manuscript. D.Y. Logunov and A.L. Gintsburg approved the final version of the manuscript.
Ethics approval. The study was approved by the Bioethics Committee at the National Research Centre for Epidemiology and Microbiology named after the honorary academician N.F.Gamaleya, Ministry of Health of the Russian Federation (Protocol No. 35 dated 24 January 2023).
Additional information. Tables 1–4 are published on the website of Biological Products. Prevention, Diagnosis, Treatment.
https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table1
https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table2
https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table3
https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table4
1. ICTV https://ictv.global/report/chapter/filoviridae/filoviridae/orthoebolavirus
2. https://www.cdc.gov/ebola/outbreaks/index.html
3. https://www.fda.gov/news-events/press-announcements/first-fda-approved-vaccine-prevention-ebola-virus-disease-marking-critical-milestone-public-health
4. ГОСТ 33044-2014. Принципы надлежащей лабораторной практики.
5. https://www.ncbi.nlm.nih.gov/genome/viruses/variation/
6. https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table1
7. https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table2
8. https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table3
9. https://www.ncbi.nlm.nih.gov/genome/viruses/variation/
10. https://doi.org/10.30895/2221-996X-2024-24-3-294-311-table4
11. Там же
1. Dolzhikova IV, Zubkova OV, Tukhvatulin AI, Dzharullaeva AS, Tukhvatulina NM, Shcheblyakov DV, et al. Safety and immunogenicity of GamEvac-Combi, a heterologous VSVand Ad5-vectored Ebola vaccine: An open phase I/II trial in healthy adults in Russia. Hum Vaccin Immunother. 2017;13(3):613–20. https://doi.org/10.1080/21645515.2016.1238535
2. Zhang Z, Zhao Z, Wang Y, Wu S, Wang B, Zhang J, et al. Comparative immunogenicity analysis of intradermal versus intramuscular immunization with a recombinant human adenovirus type 5 vaccine against Ebola virus. Front Immunol. 2022;13:963049. https://doi.org/10.3389/fimmu.2022.963049
3. Ahmed I, Salsabil L, Hossain MJ, Shahriar M, Bhuiyan MA, Islam MR. The recent outbreaks of Marburg virus disease in African countries are indicating potential threat to the global public health: Future prediction from historical data. Health Sci Reports. 2023;6(7):e1395. https://doi.org/10.1002/hsr2.1395
4. Rojas M, Monsalve DM, Pacheco Y, Acosta-Ampudia Y, Ramírez-Santana C, Ansari AA, et al. Ebola virus disease: An emerging and re-emerging viral threat. J Autoimmun. 2020;106:102375. https://doi.org/10.1016/j.jaut.2019.102375
5. Jones SM, Feldmann H, Ströher U, Geisbert JB, Fernando L, Grolla A, et al. Live attenuated recombinant vaccine pro tects nonhuman primates against Ebola and Marburg vi ruses. Nat Med. 2005;11(7):786–90. https://doi.org/10.1038/nm1258
6. Swenson DL, Wang D, Luo M, Warfield KL, Woraratanadharm J, Holman DH, et al. Vaccine to confer to nonhuman primates complete protection against multistrain Ebola and Marburg virus infections. Clin Vaccine Immunol. 2008;15(3):460–7. https://doi.org/10.1128/CVI.00431-07
7. Afolabi MO, Ishola D, Manno D, Keshinro B, Bockstal V, Rogers B, et al. Safety and immunogenicity of the twodose heterologous Ad26.ZEBOV and MVA-BN-Filo Ebola vaccine regimen in children in Sierra Leone: a ran domised, double-blind, controlled trial. Lancet Infect Dis. 2022;22(1):110–22. https://doi.org/10.1016/S1473-3099(21)00128-6
8. Peng W, Rayaprolu V, Parvate AD, Pronker MF, Hui S, Parekh D, et al. Glycan shield of the ebolavirus envelope glycoprotein GP. Commun Biol. 2022;5(1):785. https://doi.org/10.1038/s42003-022-03767-1
9. Ahmad I, Fatemi SN, Ghaheri M, Rezvani A, Khezri DA, Natami M, et al. An overview of the role of Niemann– Pick C1 (NPC1) in viral infections and inhibition of viral infections through NPC1 inhibitor. Cell Commun Signal. 2023;21(1):352. https://doi.org/10.1186/s12964-023-01376-x
10. Murin CD, Gilchuk P, Crowe JE, Ward AB. Structural biology illuminates molecular determinants of broad ebolavirus neutralization by human antibodies for pan-ebolavirus therapeutic development. Front Immunol. 2022;12:808047. https://doi.org/10.3389/fimmu.2021.808047
11. Зубкова ОВ, Ожаровская ТА, Должикова ИВ, Попова О, Щебляков ДВ, Гроусова ДМ и др. Экспрессионный вектор для создания иммунобиологического средства для индукции специфического иммунитета против вируса тяжелого острого респираторного синдрома SARS-CoV-2 (варианты). Патент Российской Федерации № 2731356 C9; 2021. EDN: KSWWVX
12. Lock M, Korn M, Wilson J, Sena-Esteves M, Gao G. Measuring the infectious titer of recombinant adenovirus using tissue culture infection dose 50% (TCID50) end-point dilution and quantitative polymerase chain reaction (qPCR). Cold Spring Harb Protoc. 2019;2019(8):pdb.prot095562. https://doi.org/10.1101/pdb.prot095562
13. Whitt MA. Generation of VSV pseudotypes using recombinant ΔG-VSV for studies on virus entry, identification of entry inhibitors, and immune responses to vaccines. J Virol Methods. 2010;169(2):365–74. https://doi.org/10.1016/j.jviromet.2010.08.006
14. Ожаровская ТА, Попова О, Зубкова ОВ, Вавилова ИВ, Почтовый АА, Щебляков ДЕ и др. Резработка и характеристика векторной системы на основе аденовируса обезьян 25-го серотипа. Вестник Российского государственного медицинского университета. 2023;(1):4–11. https://doi.org/10.24075/vrgmu.2023.006
15. Carroll SA, Towner JS, Sealy TK, McMullan LK, Khris tova ML, Burt FJ, et al. Molecular evolution of viruses of the family Filoviridae based on 97 whole-genome sequences. J Virol. 2013;87(5):2608–16. https://doi.org/10.1128/JVI.03118-12
16. Yang ZY, Duckers HJ, Sullivan NJ, Sanchez A, Nabel EG, Nabel GJ. Identification of the Ebola virus glycoprotein as the main viral determinant of vascular cell cytotoxicity and injury. Nat Med. 2000;6(8):886–9. https://doi.org/10.1038/78645
17. Zhao Y, Ren J, Harlos K, Jones DM, Zeltina A, Bowden TA, et al. Toremifene interacts with and destabilizes the Ebola virus glycoprotein. Nature. 2016;535(7610):169–72. https://doi.org/10.1038/nature18615
18. Cottingham MG, Carroll F, Morris SJ, Turner AV, Vaughan AM, Kapulu MC, et al. Preventing spontaneous genetic rearrangements in the transgene cassettes of adenovirus vec tors. Biotechnol Bioeng. 2012;109(3):719–28. https://doi.org/10.1002/bit.24342
19. Gallais Y, Sierocki R, Lhomme G, Sivelle C, Kiseljak D, Wurm F, et al. Large-scale mapping of the Ebola NP and GP proteins reveals multiple immunoprevalent and conserved CD4 T-cell epitopes. Cell Mol Immunol. 2021;18(5):1323–5. https://doi.org/10.1038/s41423-020-0455-2
20. Bhatia B, Furuyama W, Hoenen T, Feldmann H, Marzi A. Ebola virus glycoprotein domains associated with protective efficacy. Vaccines. 2021;9(6):630. https://doi.org/10.3390/vaccines9060630
21. Meyer M, Yoshida A, Ramanathan P, Saphire EO, Collins PL, Crowe JE, et al. Antibody repertoires to the same Ebola vaccine antigen are differentially affected by vaccine vectors. Cell Rep. 2018;24(7):1816–29. https://doi.org/10.1016/j.celrep.2018.07.044
22. Kimble JB, Malherbe DC, Meyer M, Gunn BM, Karim MM, Ilinykh PA, et al. Antibody-mediated protective mecha nisms induced by a trivalent parainfluenza virus-vectored ebolavirus vaccine. J Virol. 2019;93(4):e01845–18. https://doi.org/10.1128/JVI.01845-18
23. Murin CD, Fusco ML, Bornholdt ZA, Qiu X, Olinger GG, Zeitlin L, et al. Structures of protective antibodies reveal sites of vulnerability on Ebola virus. Proc Natl Acad Sci USA. 2014;111(48):17182–7. https://doi.org/10.1073/pnas.1414164111
24. Saphire EO, Schendel SL, Fusco ML, Gangavarapu K, Gunn BM, Wec AZ, et al. Systematic analysis of monoclonal antibodies against Ebola virus GP defines features that contribute to protection. Cell. 2018;174(4):938–952.e13. https://doi.org/10.1016/j.cell.2018.07.033
25. Bornholdt ZA, Turner HL, Murin CD, Li W, Sok D, Souders CA, et al. Isolation of potent neutralizing antibodies from a survivor of the 2014 Ebola virus outbreak. Science. 2016;351(6277):1078–83. https://doi.org/10.1126/science.aad5788
26. Towner JS, Sealy TK, Khristova ML, Albariño CG, Conlan S, Reeder SA, et al. Newly discovered Ebola virus associated with hemorrhagic fever outbreak in Uganda. PLoS Pathog. 2008;4(11):e1000212. https://doi.org/10.1371/journal.ppat.1000212
27. Hensley LE, Mulangu S, Asiedu C, Johnson J, Honko AN, Stanley D, et al. Demonstration of cross-protective vaccine immunity against an emerging pathogenic Ebolavirus species. PLoS Pathog. 2010;6(5):e1000904. https://doi.org/10.1371/journal.ppat.1000904
28. Hevey M, Negley D, Pushko P, Smith J, Schmaljohn A. Marburg virus vaccines based upon alphavirus replicons protect guinea pigs and nonhuman primates. Virology. 1998;251(1):28–37. https://doi.org/10.1006/viro.1998.9367
29. Zhu F-C, Guan X-H, Li Y-H, Huang J-Y, Jiang T, Hou L-H, et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: a randomised, double-blind, placebocontrolled, phase 2 trial. Lancet. 2020;396(10249):479–88. https://doi.org/10.1016/S0140-6736(20)31605-6
30. Flickinger JC, Singh J, Carlson R, Leong E, Baybutt TR, Barton J, et al. Chimeric Ad5.F35 vector evades anti-adenovirus serotype 5 neutralization opposing GUCY2C-targeted antitumor immunity. J Immunother Cancer. 2020;8(2):e001046. https://doi.org/10.1136/jitc-2020-001046
31. Flickinger JC, Staudt RE, Singh J, Carlson RD, Barton JR, Baybutt TR, et al. Chimeric adenoviral (Ad5.F35) and listeria vector prime-boost immunization is safe and effective for cancer immunotherapy. NPJ Vaccines. 2022;7(1):61. https://doi.org/10.1038/s41541-022-00483-z
32. Mitchell DAJ, Dupuy LC, Sanchez-Lockhart M, Palacios G, Back JW, Shimanovskaya K, et al. Epitope mapping of Ebola virus dominant and subdominant glycoprotein epitopes facilitates construction of an epitope-based DNA vaccine able to focus the antibody response in mice. Hum Vaccin Immunother. 2017;13(12):2883–93. https://doi.org/10.1080/21645515.2017.1347740
Ожаровская Татьяна Андреевна, канд. биол. наук
ул. Гамалеи, д. 18, Москва, 123098
Зубкова Ольга Вадимовна, канд. биол. наук
ул. Гамалеи, д. 18, Москва, 123098
Попова Ольга
ул. Гамалеи, д. 18, Москва, 123098
Ковыршина Анна Витальевна
ул. Гамалеи, д. 18, Москва, 123098
Голдовская Полина Павловна
ул. Гамалеи, д. 18, Москва, 123098
Вавилова Ирина Викторовна
ул. Гамалеи, д. 18, Москва, 123098
Должикова Инна Вадимовна, канд. биол. наук
ул. Гамалеи, д. 18, Москва, 123098
Ермолова Екатерина Ивановна, канд. биол. наук
ул. Гамалеи, д. 18, Москва, 123098
Кунда Марина Сергеевна, канд. биол. наук
ул. Гамалеи, д. 18, Москва, 123098
Рыжова Наталья Николаевна, канд. биол. наук
ул. Гамалеи, д. 18, Москва, 123098
Воронина Ольга Львовна, канд. биол. наук, доц.
ул. Гамалеи, д. 18, Москва, 123098
Щербинин Дмитрий Николаевич, канд. биол. наук
ул. Гамалеи, д. 18, Москва, 123098
Щебляков Дмитрий Викторович, канд. биол. наук
ул. Гамалеи, д. 18, Москва, 123098
Логунов Денис Юрьевич, д-р биол. наук, академик РАН
ул. Гамалеи, д. 18, Москва, 123098
Гинцбург Александр Леонидович, д-р биол. наук, академик РАН, профессор
ул. Гамалеи, д. 18, Москва, 123098

|
1. Таблица 1. Программа амплификации для постановки полимеразной цепной реакции в режиме реального времени (ПЦР-РВ) на ген гексона Ad5 | |
| Тема | ||
| Тип | Исследовательские инструменты | |
Скачать
(617KB)
|
Метаданные ▾ | |
|
|
2. Таблица 2. Нуклеотидные последовательности праймеров и зонда | |
| Тема | ||
| Тип | Исследовательские инструменты | |
Скачать
(729KB)
|
Метаданные ▾ | |
|
|
3. Таблица 3. Программа амплификации для постановки полимеразной цепной реакции (ПЦР) на целевой ген | |
| Тема | ||
| Тип | Исследовательские инструменты | |
Скачать
(702KB)
|
Метаданные ▾ | |
|
|
4. Таблица 4. Количественный и качественный анализ полученных препаратов рекомбинантных аденовирусов (rAd5) | |
| Тема | ||
| Тип | Research Instrument | |
Скачать
(571KB)
|
Метаданные ▾ | |
Ожаровская Т.А., Зубкова О.В., Попова О., Ковыршина А.В., Голдовская П.П., Вавилова И.В., Должикова И.В., Ермолова Е.И., Кунда М.С., Рыжова Н.Н., Воронина О.Л., Щербинин Д.Н., Щебляков Д.В., Логунов Д.Ю., Гинцбург А.Л. Иммуногенность различных вариантов гена гликопротеина вирусов Эбола и Марбург в составе рекомбинантных аденовирусных векторов. БИОпрепараты. Профилактика, диагностика, лечение. 2024;24(3):294-311. https://doi.org/10.30895/2221-996X-2024-24-3-294-311
Ozharovskaia T.A., Zubkova O.V., Popova O., Kovyrshina A.V., Goldovskaya P.P., Vavilova I.V., Dolzhikova I.V., Ermolova E.I., Kunda M.S., Ryzhova N.N., Voronina O.L., Shcherbinin D.N., Shcheblyakov D.V., Logunov D.Y., Gintsburg A.L. Immunogenicity of various variants of Ebola and Marburg virus glycoprotein genes in recombinant adenoviral vectors. Biological Products. Prevention, Diagnosis, Treatment. 2024;24(3):294-311. (In Russ.) https://doi.org/10.30895/2221-996X-2024-24-3-294-311

Издатель: ФГБУ «НЦЭСМП» Минздрава России































