



Contents
Scroll to:
 A. A. Nikonova,
A. S. Poroshina,
T. G. Samartseva,
A. G. Samartseva,
A. G. Adelfinskaya,
B. Tahhan,
N. N. Shershakova,
M. R. Khaitov,
A. S. Oksanich,
G. M. Ignatyev,
V. V. Zverev
A. A. Nikonova,
A. S. Poroshina,
T. G. Samartseva,
A. G. Samartseva,
A. G. Adelfinskaya,
B. Tahhan,
N. N. Shershakova,
M. R. Khaitov,
A. S. Oksanich,
G. M. Ignatyev,
V. V. Zverev https://doi.org/10.30895/2221-996X-2026-26-1-42-53
Scroll to:
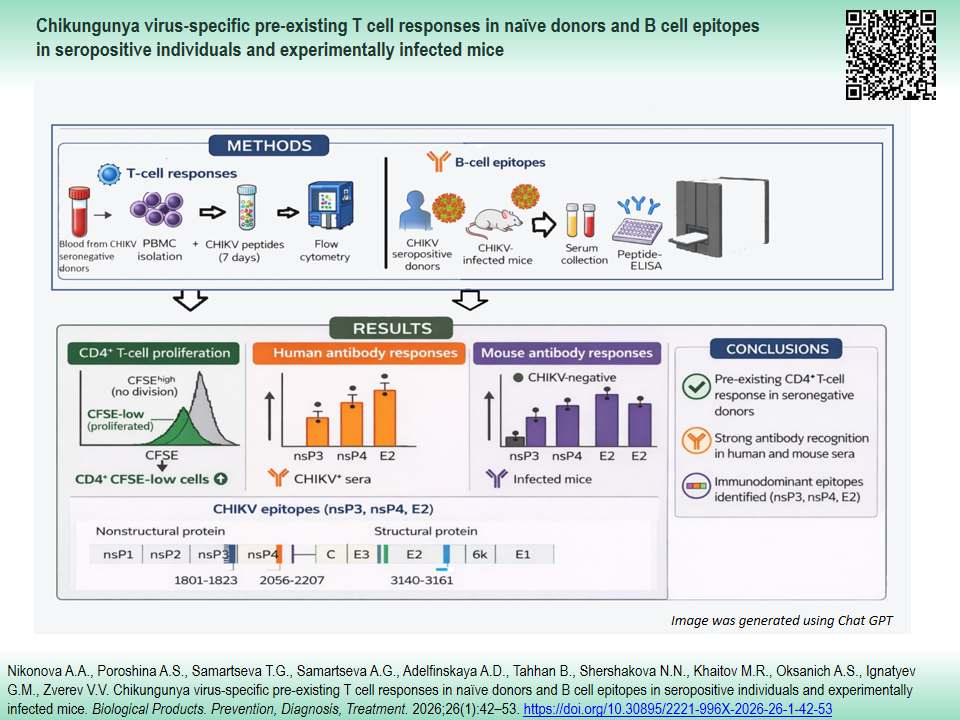
INTRODUCTION. Chikungunya virus (CHIKV) poses a major public health challenge in endemic regions due to the lack of specific preventive measures and effective antiviral therapies. Understanding and identifying viral epitopes through rigorous research directly supports the development of more effective next-generation vaccines.
AIM. This study aimed to investigate the presence of pre-existing CHIKV-specific T cell memory in unexposed individuals and identified the B cell epitopes targeted in both seropositive donors and CHIKV-infected murine models.
MATERIALS AND METHODS. Peripheral blood from 34 healthy volunteers and sera from four CHIKV-seropositive individuals, as well as from BALB/c mice immunized with a non-lethal CHIKV strain, were analyzed to assess Tand B-cell responses. Five synthetic peptides (21–29 aa) derived from nsP2, nsP3, nsP4, and E2 proteins were selected using the immunogenicity prediction algorithm from the Immune Epitope Database and synthesized to encompass predicted epitopes with flanking residues. Peripheral blood mononuclear cells from unexposed donors were stimulated with peptides (1.2 µg/mL), and CD4+/CD8+ T cell proliferation was evaluated by carboxyfluorescein succinimidyl ester (CFSE)-based flow cytometry. Human and mouse sera were tested for peptide-specific IgG by enzyme-linked immunosorbent assay (ELISA). Epitope localization was visualized on 3D protein models using UCSF ChimeraX software and web server I-TASSER.
RESULTS. T cell proliferation assays with CFSE labeling revealed preexisting CD4+, but not CD8+, T cell responses to peptides from nonstructural (nsP2–4) and structural (E2) CHIKV proteins in seronegative donors (P<0.05, P<0.01). Linear B cell epitopes within nsP3, nsP4, and E2 were identified by ELISA in sera from CHIKV-seropositive donors and CHIKV-infected mice. These epitopes were subsequently mapped onto 3D models of CHIKV proteins (E2, amino acids 3140–3161; nsP3, amino acids 1801–1823; nsP4, amino acids 2207–2256).
CONCLUSIONS. Our findings indicate the presence of preexisting CD4+ T cell responses in antigen-naïve individuals and underscore the importance of experimentally validating in silico–predicted epitopes for advancing serological diagnostics and vaccine development.
Nikonova A.A., Poroshina A.S., Samartseva T.G., Samartseva A.G., Adelfinskaya A.G., Tahhan B., Shershakova N.N., Khaitov M.R., Oksanich A.S., Ignatyev G.M., Zverev V.V. Chikungunya virus-specific pre-existing T cell responses in naïve donors and B cell epitopes in seropositive individuals and experimentally infected mice. Biological Products. Prevention, Diagnosis, Treatment. 2026;26(1):42-53. https://doi.org/10.30895/2221-996X-2026-26-1-42-53
Chikungunya virus (CHIKV), an Alphavirus of the Togaviridae family, is transmitted to humans through the bite of female Aedes mosquitoes [1]. It is an enveloped virus with a single-stranded, positive-sense RNA genome of approximately 11.8 kb. The genome contains two open reading frames that encode four nonstructural proteins (nsP1–4) required for viral replication, three major structural proteins (Capsid, E1, and E2), and two small cleavage products (E3 and 6K) [2]. The first recognized outbreak of CHIKV occurred in Tanzania in 1952–1953 [3]. Since that time, the virus has spread to more than 100 countries in tropical and subtropical regions, causing millions of reported cases globally [4]. The detection of nearly 8,000 CHIKV cases in the Southern China since June 2025 highlights the growing public health impact of this arboviral infection [5]. In humans, CHIKV initially replicates in skin fibroblasts and subsequently disseminates to the liver, muscle, joints, lymph nodes, spleen, and brain [1]. CHIKV infection causes inflammatory musculoskeletal disease in humans, characterized by debilitating symptoms such as arthralgia, arthritis, and myalgia [6].
The host response to CHIKV infection begins locally in the skin at the site of the mosquito bite, where the cutaneous immune system provides the first line of defense. During the acute phase, the virus disseminates from this initial site of replication to primary target tissues. This phase lasts for 7–14 days and is characterized by viremia, clinical symptoms, and the production of anti-CHIKV IgM antibodies [1, 7]. Active viral replication triggers the innate immune response, characterized by the production of type I interferons [1]. The early acute phase is mediated by CD8⁺ T cell responses, followed by a predominance of CD4⁺ T cells at later stages of infection [8]. The post-acute phase of CHIKV infection is characterized by viral clearance mediated by neutralizing antibodies (predominantly IgG) and elimination of infected cells by NK cells, CD8⁺ T cells, and neutrophils, leading to disease resolution [9].
Approximately 30% of individuals develop chronic disease characterized by persistent arthralgia and arthritis [1][10]. Multiple studies indicate that CHIKV can evade CD8⁺ T cell responses, which may promote chronic infection within joint tissues [11]. Due to T cell receptor (TCR) cross-reactivity, pre-existing memory phenotype T cells have been identified among human antigen-specific CD4⁺ lymphocytes, even in the absence of known viral exposure [12].
Despite significant progress in understanding humoral responses to CHIKV, the role of cellular immunity in protection and pathogenesis remains insufficiently defined. In particular, it is unclear whether pre-existing T cell memory against CHIKV exists in antigen-naïve individuals and how different viral proteins contribute to antibody recognition. This lack of knowledge limits the development of immunodiagnostic tools and rational vaccine design. We hypothesized that predicted in silico CHIKV peptides could contain epitopes capable of activating pre-existing CD4⁺ T cells in unexposed donors and inducing B cell responses in seropositive individuals and infected mice.
The aim of this study was to investigate the presence of pre-existing CHIKV-specific T cell memory responses in unexposed individuals and to identify B cell epitopes targeted in seropositive donors and CHIKV-infected murine model.
Human samples сollection. Peripheral blood samples were collected from 34 healthy volunteers (age 31±10 years; 11 women and 23 men) residing in the Moscow region. Ethics approval for both human and animal studies (Protocol No. 6, dated March 4, 2023) was obtained from the Local Research Ethics Committee of the I. Mechnikov Research Institute for Vaccines and Sera. Written informed consent was obtained from all human participants in accordance with the Declaration of Helsinki (2013 revision), the International Council for Harmonisation Good Clinical Practice (ICH-GCP) guidelines, and the national regulations of the Russian Federation, including Federal Law No. 323-FZ1, On the Fundamentals of Health Protection of Citizens in the Russian Federation.
Animals. All animal procedures, including euthanasia by cervical dislocation under light ether anesthesia, were performed in accordance with the institutional guidelines of the Mechnikov Research Institute for Vaccines and Sera, the EU Directive 2010/63/EU for animal experiments. Female BALB/c mice (6–8 weeks old; Stolbovaya, Moscow, Russia) were randomly assigned to two groups: CHIKV-infected (n=9) and PBS-treated control (n=9) (Phosphate-Buffered Saline, Cat. No. Р060Е, Paneco, Russia). Each group consisted of three independent pools (3 mice per pool). Animals were housed in a pathogen-free facility under standard conditions (22±2 °C, 50–60% humidity, 12 hours light/dark cycle) with free access to food and water. Environmental enrichment was provided with nesting material and shelters.
All manipulations, including virus inoculation and blood collection, were performed under ether anesthesia to minimize pain and distress. No animals were excluded from the study, and daily health monitoring was conducted by trained personnel.
Virus strain. The CHIKV strain Nika2021 (GenBank Acc. No. OQ320495) [13] was propagated in Aedes albopictus clone C6/36 cells (ATCC) and used for mouse immunization.
Mouse immunization. BALB/c mice were immunized intraperitoneally with a non-lethal CHIKV strain Nika2021 (100 TCID50 per mouse) twice at two-week intervals. Serum samples were collected two weeks after the final immunization. Control mice received PBS injections. Sera from three mice per group were pooled, and experiments were repeated independently three times (n=3 pools). Sera were analyzed for peptide-specific IgG by ELISA as described below.
Peptides design and synthesis. Five synthetic peptides derived from nsP2, nsP3, nsP4, and E2 proteins of CHIKV were selected from the Immune Epitope Database (IEDB) as regions with predicted high immunogenic potential [14]. The IEDB immunogenicity algorithm, trained on a large dataset of experimentally validated T cell epitopes, distinguishes immunogenic from non-immunogenic sequences by evaluating amino acid properties and their positional context within the peptide. For each viral protein, the peptide region with the highest immunogenicity score (0.18–0.26) was selected. To enable natural processing and presentation across multiple HLA alleles, and to evaluate both T and B cell responses, extended peptides of 21–29 aa encompassing the predicted core epitopes and adjacent flanking residues were synthesized. Peptides were produced by the "Human proteome" Core Facility at the Institute of Biomedical Chemistry (Moscow, Russia) (Table 1) and dissolved in dimethyl sulfoxide (DMSO) at ~1800 pmol/µL.
Table 1. Sequences of linear peptides identified by in silico analysis
Таблица 1. Последовательности линейных пептидов, идентифицированных in silico
|
Peptide name Название пептида |
Sequence Последовательность |
|
D03-NSP1 |
CPMRSAEDPERLANYARKLAS |
|
C04 -NSP2 |
TDEESYELVRAERTEHEYVYDVD |
|
D01-NSP3 |
GASSETFPITFGDFNEGEIESL |
|
C02-NSP4 |
LFDMSAEDFDAIIAAHFKPGDTVLETDIA |
|
D02-E2 |
LSTNGTAHGHPHEIILYYYEL |
T cell proliferation assay. Peripheral blood mononuclear cells (PBMCs) were isolated from healthy donors (n=12) by Ficoll-Hypaque density gradient centrifugation. Cells were resuspended in serum-free medium (Cat. No. 12-725F, UltraCULTURE™, Lonza, Switzerland) and seeded in 96-well round-bottom plates (2×10⁵ cells/well). Carboxyfluorescein succinimidyl ester, CFSE, (Cat. No. C34554, Invitrogen, USA) was added at a final concentration of 0.00002 mM. CFSE-labeled cells were stimulated with peptides at concentrations 1.2 µg/mL for 7 days. Anti-CD3/CD28 Dynabeads™ (Cat. No. 11131D, Gibco, USA) were used as positive control, and medium alone as negative control.
After culture, cells were stained with the eBioscience™ Fixable Viability Dye eFluor™ 660 (Cat. No. 65-0865-14, eBioscience, USA) and surface anti-human CD3, CD4, CD8 antibodies (Cat. No. 12-0038-42, 25-0049-42, A15448, respectively, eBioscience). Nonspecific binding was blocked with 10% normal mouse serum (Cat. No. 10410, Invitrogen, USA). Flow cytometry was performed on a BD FACSCanto II (BD Biosciences, USA), and data were analyzed using FlowJo v10.6.2 (BD Biosciences, USA). To quantify proliferation, viable CD3⁺CD4⁺ and CD3⁺CD8⁺ T cells were gated, and the proportion of CFSElow cells was determined (Figure 1).


Fig. 1. Gating strategy for analysis of CD4⁺ or CD8⁺ T cells proliferation using cell-tracking dye carboxyfluorescein succinimidyl ester (CFSE). Flow cytometry was performed on human peripheral blood mononuclear cells (n=12). The lymphocytes were gated based on the forward versus side scatter (A) and after the exclusion of debris, doublets and dead cells (A–C), CD4⁺ and CD8⁺ T cells were identified on the gated CD3⁺ cells (D, E). The intensity of CFSE was evaluated on the gated CD3⁺CD4⁺ (F, H, J, L) and CD3⁺CD8⁺ T cells (G, I, K, M). To determine T cell proliferation the percentages of CFSE positive and negative cells were determined on the gated cells. Dot plot and histogram examples showing the percentage of CFSE positive and negative CD3⁺CD4⁺ and CD3⁺CD8⁺ T cells from one of the analyzed samples (A–M) treated with Dynabeads™ (F and G, positive control), media (H and I, negative control), C04-NSP2 peptide at 1.2 µg/mL (J and K), respectively. Histograms (L, M) demonstrating CFSE unstained cells.
Рис. 1. Алгоритм анализа пролиферации CD4⁺ или CD8⁺ Т-клеток с использованием красителя сукцинимидилового эфира карбоксифлуоресцеина (CFSE). Проточную цитометрию проводили с использованием мононуклеаров периферической крови человека (n=12). Идентификация лимфоцитов проводилась с помощью диаграммы бокового — прямого (FSS–SSC) светорассеяния (А), после исключения дуплетов и мертвых клеток из анализа (A–С) идентифицировали популяции CD3⁺CD4⁺ и CD3⁺CD8⁺ Т-клеток (D, E). Интенсивность окрашивания CFSE оценивали в CD3⁺CD4⁺ (F, H, J, L) и CD3⁺CD8⁺ Т-клетках (G, I, K, M). Для оценки пролиферации Т-клеток определяли процентное соотношение CFSE-положительных и отрицательных клеток в идентифицированных лимфоцитах. На рисунке представлены примеры точечных диаграмм и гистограмм, отражающих долю CFSE-положительных и CFSE-отрицательных CD3⁺CD4⁺ и CD3⁺CD8⁺ Т-клеток в одном из анализируемых образцов (A–M): стимуляция Dynabeads™ (F, G — положительный контроль), культивирование в питательной среде (H, I — отрицательный контроль) и обработка пептидом C04-NSP2 в концентрации 1,2 мкг/мл (J, K). Гистограммы (L, M) демонстрируют клетки, не окрашенные CFSE.
Identification of CHIKV B-cell epitopes by ELISA. Five peptides (Table 1) were biotinylated using a Biotin Protein Labeling Kit (Roche, Switzerland). Streptavidin-coated plates (Biomat, Italy) were loaded with 12 pmol/well of peptide and incubated with serum samples (human or murine). Human sera included CHIKV-IgG–positive (n=4) and negative (n=18) samples confirmed by commercial ELISA (Bioservice BTC, Russia). Mouse sera were collected from CHIKV-infected or PBS-treated animals. Bound IgG was detected with HRP-conjugated anti-human or anti-mouse IgG (Bioservice BTC, Russia) and developed with TMB substrate. Absorbance was read at 450 nm with a 620 nm reference in a microplate autoreader (Erba Mannheim, Germany). A peptide was considered to contain a positive B cell epitope when OD values obtained with CHIKV-positive sera exceeded the mean +3 SD of negative sera.
Computational modeling of protein structures. Structural data for nsP4 were obtained from the Protein Data Bank (PDB ID: 7Y38) [15] and visualized with UCSF ChimeraX software [16]. Predicted 3D structures of nsP3 and E2 were generated using I-TASSER [17] and visualized with UCSF ChimeraX [16].
Statistical analysis. The Shapiro — Wilk test for normality was performed. Data were presented as means ±SD (standard deviation) for parametric analyses and as medians ± interquartile range for nonparametric analyses, as indicated in the figure legends. Statistical analyses were performed using Prism 8 (GraphPad Software). For not normally distributed data the Kruskall — Wallis test followed by post-hoc testing (if the Kruskal — Wallis was significant) using un-paired Mann — Whitney U-tests was performed, as mentioned in the respective figure legends. Data were accepted as significantly different when p-value <0.05.
Predicted CHIKV peptides (Table 1) were tested for their ability to induce T cell proliferation in PBMCs from seronegative donors. Cells were CFSE-labeled, cultured with peptides at 1.2 µg/mL for 7 days and analyzed by flow cytometry (Figure. 2A). Significant proliferation of CD4⁺ T cells was observed in response to peptides C04-NSP2, D01-NSP3, C02-NSP4, and D02-E2 (P<0.05, P<0.01) compared with unstimulated control (Figure 2A). No significant proliferation of CD8⁺ T cells was detected at either peptide concentration. As expected, stimulation with anti-CD3/CD28 Dynabeads™ (Gibco, USA) induced robust proliferation of both CD4⁺ and CD8⁺ T cells (P<0.0001).

Fig. 2. Activation of Chikungunya virus (CHIKV)-specific CD4⁺ (A) and CD8⁺ (B) T lymphocytes isolated from blood of healthy donors (n=12). A, proliferation level of CD3⁺CD4⁺ T lymphocytes; B, proliferation level of CD3⁺CD8⁺ T lymphocytes. The proliferation was evaluated by flow cytometry using carboxyfluorescein succinimidyl ester (CFSE). Peripheral blood mononuclear cells were isolated from the blood of healthy donors by Ficoll-Hypaque density gradient centrifugation and stimulated with CHIKV-derived peptides at concentrations 1.2 µg/mL for 7 days. The Dynabeads™ Human T-Activator CD3/CD28 and cells treated with media alone were used as positive and negative control, respectively. Differences between multiple groups were estimated using a Kruskal — Wallis test followed by post-hoc testing (if the Kruskal — Wallis criterion was significant) using un-paired Mann — Whitney U-tests. Horizontal bars and error bars are medians + interquartile range (12 donors). * P<0.05, ** P<0.01, **** P<0.0001. Abbreviations: nsP1, nsP2, nsP3, nsP4, nonstructural protein 1, 2, 3, 4, respectively; E2, envelope protein 2.
Рис. 2. Активация CD4⁺ (A) и CD8⁺ (B) Т-лимфоцитов, специфичных к вирусу Чикунгунья (ВЧИК), выделенных из крови здоровых доноров (n=12). А — уровень пролиферации Т-лимфоцитов CD3⁺CD4⁺; B — уровень пролиферации Т-лимфоцитов CD3⁺CD8⁺. Пролиферацию оценивали методом проточной цитометрии с использованием сукцинимидилового эфира карбоксифлуоресцеина (CFSE). Мононуклеарные клетки периферической крови выделяли из крови здоровых доноров путем центрифугирования в градиенте плотности фиколл-гипак и стимулировали специфичными пептидами к ВЧИК в концентрации 1,2 мкг/мл в течение 7 сут. В качестве положительного и отрицательного контролей использовали реагент Dynabeads™ Human T-Activator CD3/CD28 и клетки, обработанные только питательной средой, соответственно. Различия между группами оценивали с помощью критерия Краскела — Уоллиса с последующим апостериорным тестированием (если критерий Краскела — Уоллиса был значимым) с использованием непарных U-критериев Манна — Уитни. Горизонтальные полосы и планки погрешностей представляют собой медианные значения и межквартильные размахи (n=12). * P<0,05, ** P<0,01, **** P<0,0001. Сокращения: nsP1, nsP2, nsP3, nsP4 — неструктурные белки 1, 2, 3, 4, соответственно; E2 — оболочечный белок 2; media — питательная среда.
Biotinylated peptides were tested by ELISA with sera from CHIKV-seropositive and seronegative donors (Figure 3A). Three peptides D01-NSP3, C02-NSP4 and D02-E2 were consistently recognized by antibodies from CHIKV-positive individuals but not by seronegative controls. The same peptides were further analyzed using sera from CHIKV-infected mice. At both 1:100 and 1:200 dilutions, sera from infected animals showed strong reactivity to these peptides, whereas sera from PBS-treated controls did not (Figure 3B).

Fig. 3. Identification of Chikungunya virus (CHIKV) B cell epitopes. A, human blood sera from CHIKV-IgG-positive donors (n=4) and CHIKV-IgG-negative controls (n=18) were tested at 1:100 dilution in peptide-based enzyme-linked immunosorbent assay using peptides derived from structural (E2) and nonstructural (nsP1, nsP2, nsP3, nsP4) proteins. B, mouse blood sera collected from CHIKV infected animals (CHIKV+, n=3 pools of 3 mice each) or phosphate saline treated (CHIKV-, n=3 pools) were tested at dilutions of 1:100 and 1:200. Peptides were considered positive linear B-cell epitopes when optical density at 450 nm (OD450) values from CHIKV-IgG-positive samples exceeded the mean plus 3 standard deviations of negative controls (cut-off values indicated above the chart bars). Data are shown as individual values with median and interquartile range. * P<0.05. Abbreviations: nsP1, nsP2, nsP3, nsP4, nonstructural protein 1, 2, 3, 4, respectively; E2, envelope protein 2.
Рис. 3. Идентификация В-клеточных эпитопов вируса Чикунгунья (CHIKV). A — образцы сыворотки крови CHIKV-IgG-позитивных доноров (n=4) и CHIKV-IgG-негативных лиц, контроль (n=18) анализировали в разведении 1:100 методом пептид-специфического иммуноферментного анализа с использованием пептидов, соответствующих участкам структурного (E2) и неструктурных (nsP1, nsP2, nsP3, nsP4) белков CHIKV. B — образцы сыворотки крови мышей, инфицированных CHIKV (CHIKV+, n=3 пула по 3 особи в каждом), а также животных контрольной группы, обработанных фосфатно-солевым буфером (CHIKV-, n=3 пула), тестировали в разведениях 1:100 и 1:200. Пептиды расценивали как положительные линейные В-клеточные эпитопы в случае, если значения оптической плотности при длине волны 450 нм (OD450) в образцах CHIKV-IgG-позитивных сывороток превышали среднее значение отрицательных контролей более чем на три стандартных отклонения (пороговые значения указаны над столбцами). Данные представлены в виде индивидуальных значений с указанием медианы и межквартильного размаха. * P<0,05. Сокращения: nsP1, nsP2, nsP3, nsP4 — неструктурные белки 1, 2, 3, 4, соответственно; E2 — оболочечный белок 2; ВЧИК — вирус Чикунгунья.
Epitope-containing regions identified in functional assays were mapped onto the 3D structure of nsP4 (PDB ID: 7Y38) and onto I-TASSER–predicted models of nsP3 and E2 (Figure 4A–C). The epitope within the E2 protein (amino acids residues 3140–3161) was located on the protein surface (Figure 4A, D). In contrast, the regions mapped to nsP3 (amino acids residues 1801–1823) and nsP4 (amino acids residues 2207–2256) were embedded within the folded structures (Figure 4B–D).

Fig. 4. Structural localization of Chikungunya virus (CHIKV) CD4⁺ T- and B-cell epitopes. A–C, identified epitope-containing regions are mapped onto models of CHIKV E2 (A), nsP4 (B), and nsP3 (C) proteins. D, schematic representation of the CHIKV proteome indicating the positions of the identified CD4⁺ T cell and B cell epitopes. Structural mapping was performed using UCSF ChimeraX; the nsP4 structure was retrieved from the Protein Data Bank (PDB ID: 7Y38), while E2 and nsP3 models were generated using the I-TASSER server. Abbreviations: nsP1, nsP2, nsP3, nsP4, nonstructural protein 1, 2, 3, 4, respectively; C, capsid protein; E1, E2, E3, envelope protein 1, 2, 3, respectively; 6k, 6k protein.
Рис. 4. Структурная локализация CD4⁺ Т- и В-клеточных эпитопов вируса Чикунгунья (CHIKV). A–C — участки белков, содержащие идентифицированные эпитопы, картированы на 3D-модели белков E2 (A), nsP4 (B) и nsP3 (C) вируса CHIKV. D — схематическое представление протеома вируса CHIKV с указанием расположения выявленных CD4⁺ Т- и В-клеточных эпитопов. Структурное картирование выполнено с использованием программы UCSF ChimeraX; структура белка nsP4 была получена из базы данных Protein Data Bank (PDB ID: 7Y38), тогда как 3D модели белков E2 и nsP3 были построены с использованием веб-сервера I-TASSER. Сокращения: nsP1, nsP2, nsP3, nsP4 — неструктурные белки 1, 2, 3, 4, соответственно; C — белок капсида; E1, E2, E3 — оболочечный белок 1, 2, 3, соответственно; 6k — 6k белок.
In November 2023, the FDA approved the first live-attenuated CHIKV vaccine, marking an important step toward controlling this disease2. Several other vaccine candidates are in development or clinical trials, underscoring the continued global relevance of CHIKV [18]. However, these vaccines are not yet widely accessible, and detailed understanding of the immune mechanisms that contribute to protection remains limited.
A better understanding of T cell immunity and precise definition of immunogenic epitopes remain critical for vaccine design and diagnostic tools. CHIKV persistence in lymphoid and joint-associated tissues is thought to result from incomplete viral clearance despite the presence of specific antibodies [19]. Evidence also suggests that CHIKV can evade CD8⁺ T cell and B cell responses [20][21].
While in silico analyses have identified numerous candidate CHIKV B cell and T cell epitopes [22][23], only a limited number of these predictions have been experimentally validated to date [24]. In this study, we sought to evaluate preexisting T cell responses in healthy donors using linear peptides derived from CHIKV proteins (nsP1, nsP2, nsP3, nsP4, and E2). Particular attention was given to the repertoire of CD8⁺ T cells in previously unexposed individuals, given the potential role of preexisting cell-mediated immunity in facilitating viral clearance [25].
T cells with a memory phenotype can be detected even in antigen-naïve individuals, a phenomenon explained by T cell cross-reactivity and described for several viral infections [12][26][27]. Based on this concept, we tested in silico-selected CHIKV peptides ex vivo in seronegative donors to assess the presence of preexisting T cell responses. The peptides were selected using the IEDB immunogenicity prediction algorithm, which evaluates amino acid composition and positional context to distinguish immunogenic from non-immunogenic regions [14]. For each protein, we chose one of the top-scoring regions and synthesized extended 21–29 aa peptides. The use of such extended peptides is consistent with previous strategies applied in both viral immunology and cancer vaccine research, where synthetic long peptides are deliberately designed to include the predicted epitope core and additional flanking residues. These long peptides can be naturally processed by antigen-presenting cells, thereby allowing simultaneous stimulation of CD4⁺ and CD8⁺ T cell responses as well as recognition by B cells [28][29]. Although this differs from the minimal epitope approach commonly used in CD8⁺ T cell assays, it provides a physiologically relevant framework to validate in silico predictions.
Our results demonstrated that several peptides, initially predicted as CD8 epitopes, induced proliferation of CD4⁺ T cells in unexposed donors, but not CD8⁺ responses. However, given that extended peptides (21–29 aa) are not optimal for direct CD8⁺ T cell stimulation, these findings should not be interpreted as evidence of an absence of CD8⁺ responses to CHIKV. Rather, they highlight the need for further studies using minimal epitopes and HLA-matched donors to comprehensively assess the CD8⁺ repertoire.
Furthermore, biotinylated peptides were tested in ELISA with CHIKV-seropositive and seronegative human serum samples as well as sera from CHIKV-infected and uninfected mice. These experiments demonstrated that the same CHIKV-derived peptides (nsP3, nsP4, and E2) were recognized by antibodies in both humans and mice (Figure 3). Notably, the regions identified included both structural (E2 glycoprotein) and nonstructural proteins (nsP3 and nsP4), indicating that humoral responses to CHIKV are not restricted to structural antigens. These findings are consistent with the work of Yiu-Wing Kam et al., who demonstrated in experimentally infected macaques that antibody responses target multiple CHIKV proteins, including C, E2, nsP1, nsP3, and nsP4 [30].
Mapping of these epitopes onto structural models provided further biological context. The epitope within the E2 glycoprotein was surface-exposed, consistent with its role as the main structural target of the humoral response [30–32]. In contrast, the identified regions within nsP3 and nsP4 were located in buried regions of the folded proteins, which may influence their accessibility in vivo. Interestingly, the nsP3 epitope overlapped with the hypervariable C-terminal region, a domain implicated in virus–host interactions and pathogenesis [33][34].
This study has several limitations. Only a small number of peptides were tested, thus the results cannot be generalized to the entire CHIKV proteome. The use of extended 21–29 amino acid peptides differs from the typical lengths of CD8 (8–11 aa) and CD4 (≤20 aa) epitopes and may have influenced the pattern of T cell activation. The number of CHIKV-seropositive human samples was small, and larger cohorts are needed to confirm the findings. Finally, functional antibody assays were restricted to peptide binding and did not directly assess neutralization.
In summary, this study demonstrates the presence of preexisting CD4⁺ T cell responses to selected CHIKV-derived peptides in unexposed individuals and identifies linear B cell epitopes within both structural (E2) and nonstructural (nsP3 and nsP4) proteins recognized by sera from seropositive donors and infected mice. While current CHIKV vaccine strategies mainly emphasize CD8⁺ T cell-mediated protection, CD4⁺ T cells play a central role in supporting cytotoxic T lymphocyte responses, antibody maturation, and long-term immune memory. Therefore, the identification of CHIKV-specific CD4⁺ T cell and B cell epitopes may contribute to a broader understanding of protective immunity and complement CD8-focused approaches. This fact may stimulate the development of diagnostic tools and support the rational design of next-generation vaccines.
Authors’ contributions. All the authors confirm that they meet the ICMJE criteria for authorship. The most significant contributions were as follows. A.A. Nikonova conceptualized the study, drafted the manuscript, formulated the conclusions. A.S. Poroshina, T.G. Samartseva, A.G. Samartseva, A.D. Adelfinskaya, B. Tahhan, and N.N. Shershakova carried out experiments. M.R. Khaitov, A.S. Oksanich, G.M. Ignatyev, and V.V. Zverev participated in formulating the conclusions and approved the final version of the manuscript for publication.
Ethics approval. The study protocols involving human subjects and experimental animals were approved by the Local Research Ethics Committee of the I. Mechnikov Research Institute for Vaccines and Sera (Protocol No. 6 dated March 4, 2023).
Вклад авторов. Все авторы подтверждают соответствие своего авторства критериям ICMJE. Наибольший вклад распределен следующим образом: А.А. Никонова — концепция работы, написание текста рукописи, формулировка выводов; А.С. Порошина, Т.Г. Самарцева, А.Г. Самарцева, А.Д. Адельфинская, Б. Тахан, Н.Н. Шершакова — проведение экспериментов; М.Р. Хаитов, А.С. Оксанич, Г.М. Игнатьев, В.В. Зверев — участие в формулировании выводов, утверждение окончательной версии рукописи для публикации.
Соответствие принципам этики. Исследования с участием добровольцев и экспериментальных животных были одобрены Локальным комитетом по этике при ФГБНУ НИИВС им. И. И. Мечникова (протокол № 6 от 04.03.2023).
1. Federal Law No. 323-FZ “On the Fundamentals of Health Protection of Citizens in the Russian Federation”.
2. https://www.fda.gov/news-events/press-announcements/fda-approves-first-vaccine-prevent-disease-caused-chikungunya-virus
1. Schwartz O, Albert ML. Biology and pathogenesis of chikungunya virus. Nat Rev Microbiol. 2010;8(7):491–500. https://doi.org/10.1038/nrmicro2368
2. Strauss JH, Strauss EG. The alphaviruses: gene expression, replication, and evolution.Microbiol Rev. 1994;58(3):491–562. https://doi.org/10.1128/mr.58.3.491-562.1994
3. Wahid B, Ali A, Rafique S, Idrees M. Global expansion of chikungunya virus: mapping the 64-year history. Int J Infect Dis. 2017;58:69–76. https://doi.org/10.1016/j.ijid.2017.03.006
4. Costa LB, Barreto FKA, Barreto MCA, Santos T, et al. Epidemiology and economic burden of Chikungunya: A systematic literature review. Trop Med Infect Dis. 2023;8(6):301. https://doi.org/10.3390/tropicalmed8060301
5. Murphy F. China battles to control Chikungunya virus as 8000 cases are reported. BMJ. 2025;390:r1699. https://doi.org/10.1136/bmj.r1699
6. De Lima Cavalcanti TYV, Pereira MR, De Paula SO, Franca RFO. A review on Chikungunya virus epidemiology, pathogenesis and current vaccine development. Viruses. 2022;14(5):969. https://doi.org/10.3390/v14050969
7. Jin J, Galaz-Montoya JG, Sherman MB, et al. Neutralizing antibodies inhibit Chikungunya virus budding at the plasma membrane. Cell Host Microbe. 2018;24(3):417–28.e5. https://doi.org/10.1016/j.chom.2018.07.018
8. Wauquier N, Becquart P, Nkoghe D, et al. The acute phase of Chikungunya virus infection in humans is associated with strong innate immunity and T CD8 cell activation. J Infect Dis. 2011;204(1):115–23. https://doi.org/10.1093/infdis/jiq006
9. Srivastava P, Kumar A, Hasan A, et al. Disease resolution in Chikungunya — What decides the outcome? Front Immunol. 2020;11:695. https://doi.org/10.3389/fimmu.2020.00695
10. Chopra A, Anuradha V, Ghorpade R, Saluja M. Acute Chikungunya and persistent musculoskeletal pain following the 2006 Indian epidemic: a 2-year prospective rural community study. Epidemiol Infect. 2012;140(5):842–50. https://doi.org/10.1017/S0950268811001300
11. Ware BC, Parks MG, Da Silva MOL, Morrison TE. Chikungunya virus infection disrupts MHC–I antigen presentation via nonstructural protein 2. PLoS Pathog. 2024;20(3):e1011794. https://doi.org/10.1371/journal.ppat.1011794
12. Su LF, Davis MM. Antiviral memory phenotype T cells in unexposed adults. Immunol Rev. 2013;255(1):95–109. https://doi.org/10.1111/imr.12095
13. Ignatyev GM, Oksanich AS, Kazakova EV, et al. Isolation and genetic analysis of the chikungunya virus from Aedes aegypti and Aedes albopictus mosquitoes captured in Central America. Journal of Microbiology, Epidemiology and Immunobiology. 2023;100(5):310–8 (In Russ.). EDN: UWGUML
14. Calis JJ, Maybeno M, Greenbaum JA, et al. Properties of MHC class I presented peptides that enhance immunogenicity. PLoS Comput Biol. 2013;9(10):e1003266. https://doi.org/10.1371/journal.pcbi.1003266
15. Tan YB, Chmielewski D, Law MCY, et al. Molecular architecture of the Chikungunya virus replication complex. Sci Adv. 2022;8(48):eadd2536. https://doi.org/10.1126/sciadv.add2536
16. Pettersen EF, Goddard TD, Huang CC, et al. UCSF Chimera — A visualization system for exploratory research and analysis. J Comput Chem. 2004;25(13):1605–12. https://doi.org/10.1002/jcc.20084
17. Zhang Y. I-TASSER server for protein 3D structure prediction. BMC Bioinformatics. 2008;9:40. https://doi.org/10.1186/1471-2105-9-40
18. Shaw CA, August A, Bart S, et al. A phase 1, randomized, placebo-controlled, dose-ranging study to evaluate the safety and immunogenicity of an mRNA-based Chikungunya virus vaccine in healthy adults. Vaccine. 2023;41(26):3898–906. https://doi.org/10.1016/j.vaccine.2023.04.064
19. Hawman DW, Stoermer KA, Montgomery SA, et al. Chronic joint disease caused by persistent Chikungunya virus infection is controlled by the adaptive immune response. J Virol. 2013;87(24):13878–88. https://doi.org/10.1128/JVI.02666-13
20. Davenport BJ, Bullock C, Mccarthy MK, et al. Chikungunya virus evades antiviral CD8(+) T cell responses to establish persistent infection in joint-associated tissues. J Virol. 2020;94(9):e02036-19. https://doi.org/10.1128/JVI.02036-19
21. Hawman DW, Fox JM, Ashbrook AW, et al. Pathogenic Chikungunya virus evades B cell responses to establish persistence. Cell Rep. 2016;16(5):1326–38. https://doi.org/10.1016/j.celrep.2016.06.076
22. Poh CM, Chan YH, Ng LFP. Role of T cells in Chikungunya virus infection and utilizing their potential in anti-viral immunity. Front Immunol. 2020;11:287. https://doi.org/10.3389/fimmu.2020.00287
23. Venkatesan A, Chouhan U, Suryawanshi SK, Choudhari JK. An in silico approach for prediction of B cell and T cell epitope candidates against Chikungunya virus. Immunol Med. 2023;46(4): 163–74. https://doi.org/10.1080/25785826.2023.2202038
24. Lorente E, Barriga A, Garcia-Arriaza J, et al. Complex antigen presentation pathway for an HLA-A*0201-restricted epitope from Chikungunya 6K protein. PLoS Negl Trop Dis. 2017; 11(10):e0006036. https://doi.org/10.1371/journal.pntd.0006036
25. Swadling L, Diniz MO, Schmidt NM, et al. Pre-existing polymerase-specific T cells expand in abortive seronegative SARS–CoV–2. Nature. 2022;601(7891):110–7. https://doi.org/10.1038/s41586-021-04186-8
26. Pothast CR, Dijkland RC, Thaler M, Hagedoorn RS, et al. SARS-CoV-2-specific CD4(+) and CD8(+) T cell responses can originate from cross-reactive CMV-specific T cells. Elife. 2022;11:e82050. https://doi.org/10.7554/eLife.82050
27. Elias G, Meysman P, Bartholomeus E, et al. Preexisting memory CD4 T cells in naive individuals confer robust immunity upon hepatitis B vaccination. Elife. 2022;11:e68388. https://doi.org/10.7554/eLife.68388
28. Rosalia RA, Quakkelaar ED, Redeker A, et al. Dendritic cells process synthetic long peptides better than whole protein, improving antigen presentation and T–cell activation. Eur J Immunol. 2013;43(10):2554–65. https://doi.org/10.1002/eji.201343324
29. Slingluff CL Jr. The present and future of peptide vaccines for cancer: single or multiple, long or short, alone or in combination? Cancer J. 2011;17(5):343–50. https://doi.org/10.1097/PPO.0b013e318233e5b2
30. Kam YW, Lee WW, Simarmata D, et al. Unique epitopes recognized by antibodies induced in Chikungunya virus-infected non-human primates: implications for the study of immunopathology and vaccine development. PLoS One. 2014;9(4): e95647. https://doi.org/10.1371/journal.pone.0095647
31. Kam YW, Lee WW, Simarmata D, et al. Longitudinal analysis of the human antibody response to Chikungunya virus infection: implications for serodiagnosis and vaccine development. J Virol. 2012;86(23):13005–15. https://doi.org/10.1128/JVI.01780-12
32. Silva J, Cunha MDP, Pour SZ, et al. Chikungunya virus E2 structural protein B-cell epitopes analysis. Viruses. 2022;14(8):1839. https://doi.org/10.3390/v14081839
33. Foy NJ, Akhrymuk M, Shustov AV, et al. Hypervariable domain of nonstructural protein nsP3 of Venezuelan equine encephalitis virus determines cell-specific mode of virus replication. J Virol. 2013;87(13):7569–84. https://doi.org/10.1128/JVI.00720-13
34. Foy NJ, Akhrymuk M, Akhrymuk I, et al. Hypervariable domains of nsP3 proteins of New World and Old World alphaviruses mediate formation of distinct, virus-specific protein complexes. J Virol. 2013;87(4):1997–2010. https://doi.org/10.1128/jvi.02853-12
Aleksandra A. Nikonova, Cand. Sci. (Biol.)
5A Maly Kazenny Ln., Moscow 105064
Alina S. Poroshina
24 Kashirskoe Hwy, Moscow 115522
Tatiana G. Samartseva
5A Maly Kazenny Ln., Moscow 105064
Anastasia G. Samartseva
5A Maly Kazenny Ln., Moscow 105064
Anastasia D. Adelfinskaya
5A Maly Kazenny Ln., Moscow 105064
Bana Tahhan
78 Vernadsky Ave, Moscow 119454
Nadezda N. Shershakova, Cand. Sci. (Biol.)
24 Kashirskoe Hwy, Moscow 115522
Musa R. Khaitov, Dr. Sci. (Med.), Prof., Acad. RAS
24 Kashirskoe Hwy, Moscow 115522
Alexey S. Oksanich, Cand. Sci. (Biol.)
5A Maly Kazenny Ln., Moscow 105064,
78 Vernadsky Ave, Moscow 119454
George M. Ignatyev, Dr. Sci. (Med.), Prof.
5A Maly Kazenny Ln., Moscow 105064
Vitaly V. Zverev, Dr. Sci. (Biol.), Prof., Acad. RAS
5A Maly Kazenny Ln., Moscow 105064
Nikonova A.A., Poroshina A.S., Samartseva T.G., Samartseva A.G., Adelfinskaya A.G., Tahhan B., Shershakova N.N., Khaitov M.R., Oksanich A.S., Ignatyev G.M., Zverev V.V. Chikungunya virus-specific pre-existing T cell responses in naïve donors and B cell epitopes in seropositive individuals and experimentally infected mice. Biological Products. Prevention, Diagnosis, Treatment. 2026;26(1):42-53. https://doi.org/10.30895/2221-996X-2026-26-1-42-53

Publisher: Scientific Centre for Expert Evaluation of Medicinal Products































